- DOI: 10.31509/2658-607x-202584-1781
SOIL ORGANOPEDOGENESIS AS AN UNHEEDED BASIC SOIL FORMING PROCESS
![]()
© 2025 O. G. Chertov, N. V. Lukina
Isaev Centre for Forest Ecology and Productivity of the RAS
Profsoyuznaya st. 84/32 bldg. 14, Moscow, 117997, Russia
E-mail: ochertov@rambler.ru; nvl07@yandex.ru
Received: 21.05.2025
Revised: 01.07.2025
Accepted: 15.07.2025
A concept of soil organopedogenesis has been proposed as an unheeded active process of general pedogenesis reflecting the predominant significance of the biological factor in soil formation according to Vernadsky’s theory on organisms’ leading role in all natural processes in the continental part of the biosphere. A methodology is presented to reflect the role of organopedogenesis in the existing soil classifications by including in their taxonomy the types of accumulation and transformation of soil organic matter and the types of organoprofiles. The inclusion of these components will make the classification dynamic and increase its practical importance. That is especially significant for decision making at the time being given the fast negative global environmental changes.
Keywords: biological factor of soil formation, organoprofile, SOM accumulation and transformation, humus forms
The idea of the far-reaching and active role of the biological factor in the soil formation and development has been for many years close to the followers of Vernadsky’s biosphere theory (Rodin, Bazilevich, 1965; Budyko, 1984; Kovda, 1985; Odum, 1986; Glazovskaya, 1988; Gorshkov, 1995). However, to date, this role has not been analysed on the basis of new data and has not been summarised in terms of its contribution to pedogenesis (Evolyuciya pochv …, 2015; Evolyuciya, funkcionirovanie …, 2020). The below paper[1] discusses specifically the importance of the biological factor in soil genesis and especially the classification. V. I. Vernadsky’s biosphere concept (1926, 1994) serves as the theoretical foundation for scientifically sound environmental management, which is well known and accepted by the scientific community. It followed the formation of soil science as an independent earth science. The biosphere is a thin shell of our planet with active «living matter» (biomass of organism communities), which is a powerful factor in the evolution of the Earth’s crust over geological times and an active reactor in which modern biogeochemical processes take place on a planetary scale in the surface layer of the atmosphere, on the surface of the lithosphere and in the hydrosphere. At the same time, the biosphere has its own feedback mechanisms with these three spheres and ways to restore its own structure and functions after disturbances. The pedosphere is a specific active boundary layer in the structure of the biosphere between the living organisms and the continental part of the lithosphere; that is, it is the soil cover of the planet Earth (Karpachevskij, 1981; Karpachevskij et al., 1984; Vernadsky, 1994; Dobrovol’skij et al., 2010). It is a thin film where the living organisms and their metabolic products have an impact on the upper zone of weathering of rocks the Earth’s crust, ensuring the effective functioning of biosphere organisms due to the biotic circulation of matter and biogenic energy flows. It has a buffer capacity that maintains the stability of biological communities and the soil itself. The presence of the active soil biota and its waste products in the pedosphere forms a clear dividing line between the soil and the lithosphere (Shoba, 1988; Dobrovol’skij, 2009; Bahmet, 2015; Targul’yan, 2019).
Modern soil science is based on the ideas of V. V. Dokuchaev (Dokuchaev, 1883; Sibircev, 1899; Yarilov, 1904) on five factors of soil formation (climate, relief, soil-forming rocks, organisms, and time), to which the anthropogenic factor has now been added. Notably, the equivalence of action of these factors was initially postulated. This basic concept is still recognised as the basis of genetical pedology. However, already P. A. Kostychev (1886) emphasised the leading role of vegetation in soil formation, which did not fit into the principle of equal influence of soil formation factors. Presumably, the validity of these postulates depends on the time periods that one handles: on the scale of millennia, the classical definition is valid, whereas with regard to intervals of several decades and centuries, during which the soil-forming rocks and relief do not change, the point of view of P. A. Kostychev is quite justified: he emphasises the contribution of vegetation as a representative of the most dynamic biotic factor of soil formation, although the power of its influence certainly depends on other components of pedogenesis as well. Since our planet is currently experiencing rapid climate change, this factor is also dynamic, and the impact of modern climate change on soils, which has been most pronounced since the 1980s, is also manifested through the biological factor, primarily vegetation and soil biota (Lukina, 2025).
Consideration of soil formation in combination not only with the primary ecogenetic vegetation succession [endoecogenesis according to V. N. Sukachev (Sukachev, Dylis, 1964)], but also with a variety of secondary successions of forest vegetation caused by various disturbances (cutting, fires, storms with the formation of pit-and-mound topography, industrial pollution, and agricultural use), allows us to conclude that both in primary and secondary successions, accumulation of soil organic matter (SOM) is a clear trend (Duchaufour, 1979; Chertov, 1981, 1990; Karpachevskij et al., 1984; Odum, 1986; Razumovskij, 1999; Lukina et al., 2010, 2019; Danilov et al., 2024; Emmer, 1995; Nadporozhskaya et al., 2006; Kalinina et al., 2009). In the intact old-growth forests remaining on small areas (Lukina et al., 2023), which were formed during the continuous turnover of generations of key edificator species (Smirnova, 1998; Smirnova, Toropova, 2008, 2016) and in which natural processes prevailed for a long time (hundreds of years) and stands with absolutely uneven-aged structure was formed (Dyrenkov, 1984), one can expect the highest level of SOM accumulation. Interestingly, the process of SOM accumulation is a nonlinear trend with pronounced peaks and falls due to the changes in the ratio of organic matter accumulation and loss caused by carbon dioxide emissions from soils and the removal of organic matter with soil waters during natural disturbances.
Moreover, the general dynamic trend of natural soil formation is a consistent soil eutrophication with the accumulation of organiccompaunds of carbon and nitrogen, but also other biophilic elements (Chertov, Razumovskij, 1980; Chertov, 1990). Over centuries of successional vegetation changes without disturbances, this process can be seen even in podzols and peat soils (Chertov, 1981, 1990). The implication is that the «theory of a single soil-forming (sod) process» proposed by V. R. Williams (cited by Williams, 2025), after all, reflects the general trend of soil eutrophication during ecogenetic successions.
It must be admitted that if we accept the dominance of the biological factor as fundamental in the soil formation in line with Vernadsky’s biospheric ideas (Ponomareva, 1964; Rodin, Bazilevich, 1965; Chertov, Razumovskij, 1980, 1981; Karpachevskij et al., 1984; Lukina et al., 2010, 2022; Lukina et al., 2019), then SOM accumulation is a basic system-forming process that distinguishes soils from geological formations in terrestrial and shallow-water landscapes of different levels (biomes, ecosystems/biogeocenoses, ecotopes, etc.). The presence of even the smallest amount of soil (not just any) organic matter is a sign of the soil at different stages of its development. The absence of SOM is a sign of a geological or anthropogenically modified substrate or rock, but not the soil. The presence of SOM in the absence of mineral material indicates that this is soil, yet on organogenic soil-forming rocks that were originally formed both on the mineral matrix and in the aquatic environment. On the other hand, even on massive rocks, a soil film with cryptomycotic SOM and nano-processes of its accumulation can form, which then lead to the emergence of lichens (Chertov et al., 2004; Lessovaia et al., 2008) and the development of soil formation processes during primary ecogenetic succession.
It should be emphasised that SOM is the main solid-phase morphologically clearly defined macro-product of biotic influence on soil formation, and it has been studied extensively and in detail (Trofimov et al., 1986; Orlov et al., 1996; Chukov, 1997; Semenov, Kogut, 2015; Piccolo et al., 2019; and many others). Unfortunately, however, theoretical studies of SOM constitution, structure and function at the beginning of the 21st century go back to the positions of a century ago, essentially abandoning the scientific legacy of the middle of the 20th century (Semenov, Kogut, 2015; Lehmann, Kleber, 2015). SOM fractionation has been actually reduced to two physical fractions: particulate organic matter (POM) and mineral-associated organic matter (MAOM) (Semenov et al., 2023), and ignoring of the past scientific findings goes so far as to abandon the term «humus» (Lehmann, Kleber, 2015). The latter is an extremity, which, at his time, S. A. Waksman (1937) did not take the liberty to go to: dark-coloured organic matter in the upper horizons of soils with the historical term «humus» is an objective reality, no matter how our ideas about its chemical composition and physical structure change. On the one hand, the outstanding role of the «living matter of the biosphere» in the formation of SOM from green plants and being processed by soil biota as a large-scale product in the soil formation is recognised almost unanimously (Ponomareva, 1964; Karpachevskij et al., 1981; Vernadsky, 1994; Dobrovolskij et al., 2010; Semenov, Kogut, 2015; Chertov, 2025; and others). On the other hand, at the classification level, there is an amorphous position regarding the itemization of the role of SOM in pedogenesis: in the genetic classifications of soils, there is no tendency to reflect the diversity of morphological forms of SOM. Even after in the 1930s V. R. Williams (cited by Williams, 2025) declared the «dialectically justified» concept of the biological evolution of soil, there was no significant shift in the Soviet soil classification (except that the «sod-podzolic soils» subtype was added to the classification).
Existing soil classifications
In the existing soil classifications, the integral role of SOM in the soil genesis is reflected very poorly. For example, the World Reference Base for Soil Resources WRB (Mirovaya …, 2024; IUSS Working Group WRB, 2022), shows the following:
1) Soils are named based on their dominant feature (often simply by colour), with the mandatory use of only stable parameters of mineral horizons, since «the properties of the upper horizon can change rapidly over time, therefore, in the WRB system they are used for diagnostics only in rare cases» (Mirovaya …, 2024, p. 25). That is, the properties of the soil that reflect it as a dynamically developing system are considered to interfere with classification.
2) The names of the «reference soil groups» show a very distinct «abiological» classification in terms of reflecting the properties of SOM as a product of plant debris transformation by the entire soil biota. Thus, of the 32 names of the «reference soil groups» («divisions» in the Russian classification), only four are in correlation with SOM (Chernozems, Phaeozems, Umbrisols and organic soils). Moreover, in the 294 «qualifiers» (genetic traits in the form of clarifying adjectives to the name of the soil), only 14 are in correlation with SOM. This would be correct if it were not about the soil, but about the classification of weathering forms of Quaternary sediments and rocks.
In contrast to the European approach, the Russian soil classification (Klassifikaciya i diagnostika …, 2004) has a more biological and ecological approach: in 52 diagnostic horizons, 24 (46%!) are based on the SOM characteristics (Polevoj opredelitel’ …, 2008). However, in general, these soil classifications do not tend to necessary taxonomic reflect in all taxa the two basic solid phases of the soil system, i. e. soil organic matter as a product of soil formation, and the mineral basis of the entire soil. V. I. Vernadsky (1994) defined the soil as a «bio-inert body», i.e. two-component, consisting of an active biological and an inert (in the soft meaning of the word) abiogenic part, which refers to the mineral matrix. However, until now, the «inert» component dominates in soil classifications.
Despite the fact that the idea of reflecting the role of humus in classifications has been plain to see for more than a hundred years, SOM turned out to be «stepchild» of existing genetic classifications with a modest place in the general theory of pedogenesis (Sokolov, 1997; Targul’yan, 2019).
Organopedogenesis
In view of the above theoretical positions, it is proposed to introduce the concept of «organopedogenesis»[2] as the critical system-forming process of soil formation and evolution. Through SOM accumulation and transformation by autotrophic and heterotrophic organisms, it affects all functions and the overall genesis of the soil system – needless to say, taking into account and under the influence of all other classical factors of soil formation. This is true both in relation to the traditionally understood genesis with the evolution of the entire soil profile and when we are dealing with continuous processes: geochemical, hydrological, and, most of all, actual biological processes. They determine the feedback from the soil to the entire biota in the biosphere, taking into account biodiversity, productivity of natural and anthropogenic phyto-, zoo- and microbiocoenoses and the role of soil in the global carbon cycle. In other words, organopedogenesis is a multicomponent basic soil process of organnic and humus–accumulative horizons formation and the entire organoprofile of the soil. However, until now, organopedogenesis is actually an absent facet in the doctrine of soil genesis, unheeded in soil genetic classifications. Semantically, the idea of organopedogenesis is already present in genetical pedology, for example, in the term organo-«genic» soils. What are the prospects for the implementation and development of the concept of organopedogenesis?
Components of organopedogenesis
Based on the above positions, it seems practical to distinguish two morphologically pronounced results of organopedogenesis: a) different types of SOM accumulation and transformation, b) different types of organoprofiles of the entire soil system. At the same time, one can see that there is a fairly well-developed scientific basis for the development of these aspects. Firstly, there is an approach in forest pedology that fits the concept of organopedogenesis perfectly. It is the classification and diagnosis of «humus forms/types» based on morphologically pronounced signs of SOM accumulation and transformation in organic and humus-accumulative horizons, which appeared first in the West (Müller, 1887; Kubiena, 1953; Wilde, 1958; Duchaufour, 1965) and then was consistently developed in Germany (Broll et al., 2006; Wachendorf et al., 2023), the USA and Canada (Wilde, 1958; Green et al., 1993; Klinka, 1997), has been recently studied intensively in the European Union (Ponge, 2003; Humusica …, 2018; Kõlli, Köster, 2018; Zanella et al., 2018, 2022) and developed successfully in Russia (Kruedener, 1916; Tyurin, Ponomaryova, 1940; Blagovidov, Burkov, 1959; Chertov, 1974, 1981, 2025; Chertov, Nadporozhskaya, 2017, 2018). Secondly, it is the concept of soil organoprofiles (Grishina, 1986; Shoba, 1988; Min’kovskij, Shoba, 1994, 1995; Bahmet, 2015).
Types of SOM accumulation and transformation (humus forms)
The systematics and diagnostics of humus forms leans on three pillars: «raw humus» («Rohhumus, mor») with a developed litter, «soft humus» («mull») with a developed humus horizon with a shallow litter, and «moder», the transitional type which has both horizons. The term mor, back in the day was criticised in the USA by Wilde (1958) due to its negative meaning in the Russian language. This triad can also be seen in the classification of humus forms of organogenic soils according to the moisture gradient up to three types of peat bogs, and it also has varieties in the soils of meadow and steppe ecosystems (Humusica …, 2018). The Russian classification of humus forms includes a large number of transitional types (Fig. 1). As far back as in 1940, I. V. Tyurin and V. V. Ponomaryova (1940) added adjectives to the word «soil» to designate humus forms/types, which later became a widespread approach in the North-West of the Russian Federation (Blagovidov, Burkov, 1959; Chertov, 1966, 1981; Chertov et al., 1974, 1978). All these humus forms/types differ in morphological, physical and chemical parameters, first of all due to the quality of litter entering the soil: the aboveground and root litter of conifers with a low content of nitrogen and other nutrients as well as with high concentration of secondary metabolites (lignin, phenols) contribute to the formation of slowly decomposing raw humus, whereas the higher quality litter of deciduous trees that has more high content of these nutrients and lower concentrations of secondary metabolites (Lukina et al., 2008, 2019, 2021; Bioraznoobrazie …, 2021) is quickly processed by the soil fauna and microorganisms forming the mull humus form. Therefore, humus forms are clearly related to the nature and intensity of humification, as well as to the composition, productivity and biodiversity of forest vegetation (Chertov, 1981; Fedorchuk et al., 2005; Chertov, Nadporozhskaya, 2018). In turn, the diverse composition of vegetation with different quality of litter, that is, with different contents of various compounds of nitrogen, phosphorus, calcium, trace elements as well as secondary metabolites, drives the formation of various humus forms.
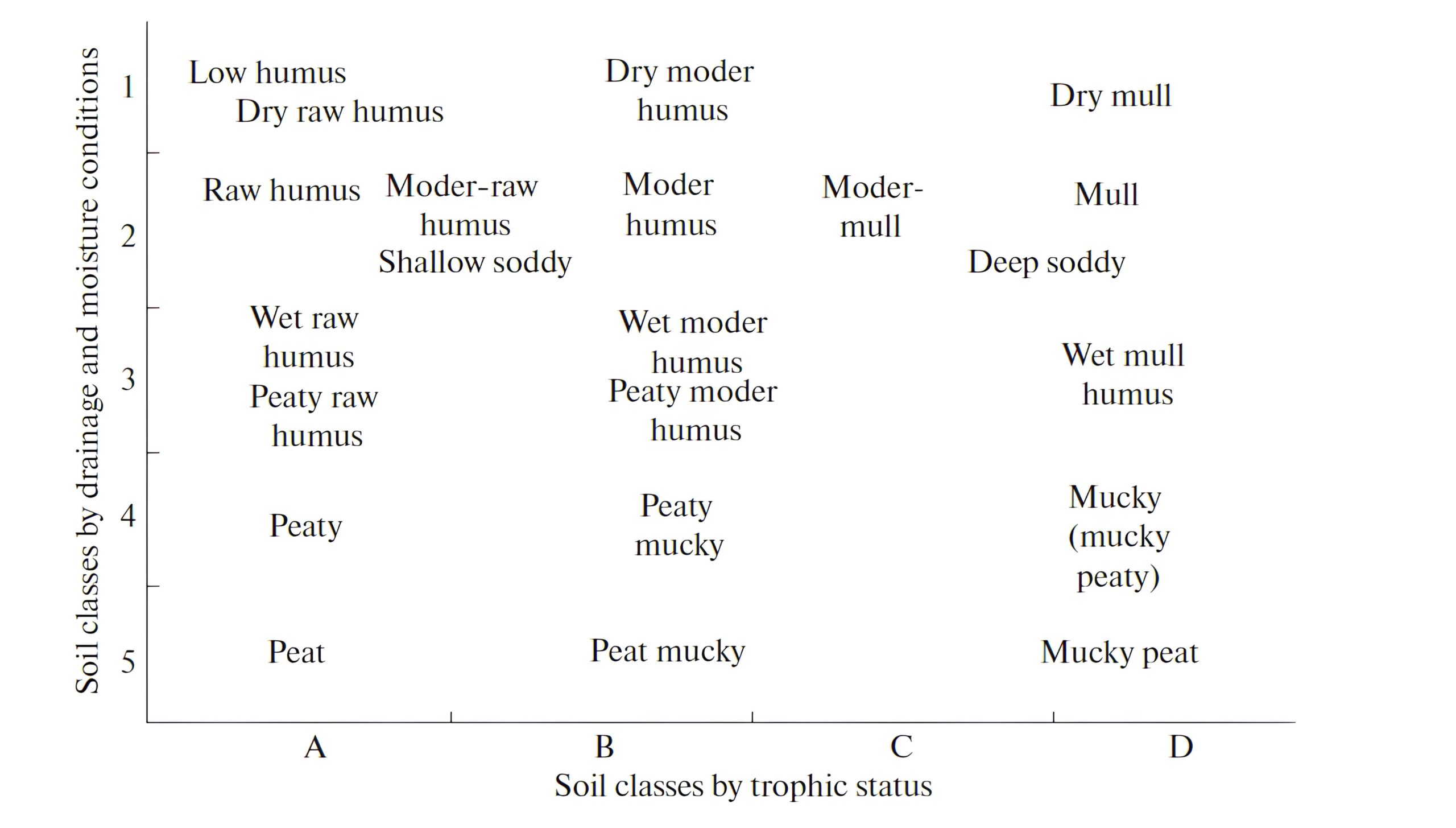
Figure 1. Ordination of types of SOM accumulation (humus forms) in forest soils by classes of richness (from A, poor to D, rich) and classes of drainage and moisture (from 1, dry to 5, constantly waterlogged) of the types of forest sites in the forest zone of European Russia (Chertov, Nadporozhskaya, 2018).
Classes of edaphic conditions are gradations of soil richness (fertility) accepted in forestry; raw humus is a synonym of mor.
This classification developed within the framework of forest pedology, but was never accepted in the Russian genetical pedology. Reasons for this may be the following: a) the concept of humus forms developed during the period of intensive dissemination of ideas of V. V. Dokuchaev on soil formation and was pushed into the category of applied forest pedology; b) there was a mistake in the translation: the German term for the Humus Form (for horizons O and Ah) was translated as a form of forest litter (Germ. Humusdecke), not humus forms. This led to the development of litter typology in the Russian soil science (Shumakov, 1958; Karpachevskij, 1981; Bogatyryov et al., 2004), and substantial investigation of forest litter has always been a part of the agenda in forest ecology and soil science. The morphological classification of humus forms has evolved for more than a century in parallel with the mainstream of genetical pedology.
The theoretical basis for identifying the types of SOM accumulation and transformation as humus forms is the now-forgotten «edaphology» (Chertov et al., 2018), which triggered the formation of soil science: in historical terms, it comes second after «agrogeology» and before the principles of genetical pedology by V. V. Dokuchaev. In edaphology, in the understanding of the Russian school, soil is considered as an underground habitat for auto- and heterotrophic organisms, primarily plant roots. Currently, this approach is closest to ecological pedology, forestry and landscape science. Please note that all existing mathematical models of SOM dynamics are edaphological (Komarov et al., 2017).
In Russia, there is a proven track record in including humus forms in the genetic classification of soils when mapping forest soils and in forest typological studies (Chertov et al., 1978; Chertov, 1981; Dyrenkov, 1984; Fedorchuk et al., 2005). In total, humus forms were included in the soil systematics on maps with a scale from 1:10,000 to 1:100,000 of seven forestry enterprises with an area of 350 thousand hectares (Chertov, 1981).
In the West, particular interest in the classification of humus forms was seen at the beginning of the 21st century due to the systematisation of available data and it even led to the replacement of the traditional term «humus form» with humipedon with the call for the development of «biological soil science» (Humusica …, 2018) since attempts to integrate them into the WRB genetic classification were unsuccessful, with the exception of France, where humus forms were included in the soil taxonomy (Duchaufour , 1979). In Western countries, classifications and field guides of humus forms are included in soil mapping manuals and forest management instructions (Malysheva et al., 2022), which was also done in Russia (Chertov, 2025).
Soil organoprofiles
The humus forms represent a classification of only the types of SOM accumulation and transformation in the area of the active biotic cycle, but not the entire soil profile (Chertov, Nadporozhskaya, 2018; Zanella et al., 2022). The soil organoprofile, as defined by the authors, is another concept referring to «the regular combination and distribution of SOM throughout the entire soil genetic profile with characteristics of its specific features» (Grishina, Orlov, 1978; Grishina, 1986). This concept was focused on a more comprehensive diagnosis of soils from the standpoint of genetical pedology. From an ecological perspective, the concept of organoprofiles was used by O. N. Bahmet (2015), who transformed it to expand the types of SOM accumulation(humus forms). However, this is not completely in line the definition of L. A. Grishina, as the same types of SOM accumulation (humus forms) can be formed in soils with different humus profiles. For example, the raw humus form can occur in soils with a monotonous SOM decrease along the profile, in soils with humus-illuvial (alfehumus) and even with a shallow undifferentiated profile on a badrock. According to F. Duchaufour (1979), these are the so-called «soil analogues».
The complete soil organoprofile reflects the release of SOM from the area of its accumulation and biotic circulation (humus forms) into the zone of biogeochemical cycles with transit, removal, eluviation and with synchronous transformation of the mineral matrix (Perelman, 1975; Fokin, 1975; Glazovskaya, 1988) caused by the influence of the biological factor. That is, according to L. A. Grishina (1986), the organoprofile is primarily a genetic, not an edaphological-ecological, concept. In the theory of soil morphogenesis by S. A. Shoba (1988), the typology of soil profiles has not been developed: the author mainly systematises the micromorphological features of the transformation processes of the mineral matrix.
Below is the authors’ draft of the systematics of morphofunctional types of organoprofiles. The authors believe that each organoprofile corresponds to a group of soil types. Therefore, there are no links to publications below for most organoprofiles, with the exception of some specific types.
Classes of organopedogenesis
- Surface-accumulative organopedogenesis (the increase in SOM upward from the surface of the mineral matrix: forest litter and turf-peat (horizons O and T); further genesis towards the formation of organogenic rocks: brown coal — hard coal (Shumakov, 1958); the extreme case is the hyper–accumulation of thick raw humus in high-altitude moist mountain forest in the tropics and the equatorial zone (Fridland, 1964; Chertov, 1990) and also “dry peat” on Russian North (Klassifikaciya i diagnostika.., 2004).
- Profile-accumulative organopedogenesis: SOM accumulation in humus-accumulative mineral horizons OA, Ah, AE and migration lower in profile in all varieties of horizons E, B, BC and C.
Types of organoprofiles in class B:
- undifferentiated;
- accumulative (monotonous, horizons A, B, C);
- hyper-accumulative – chernozem;
- agro-accumulative (all profiles with cultivated Ap, as well as Russian and German «Plaggens» with Ap thickness up to 1 m (Grigor’ev, 1980; Kalinina et al., 2009);
- eluvial and illuvial-eluvial (AE, E, EB, B);
- alfehumus (A, E, Bfh, B);
- hyper-eluvial: Ah with intensive migration of SOM in tropical rainforest (Fridland, 1964; Chertov, 1985);
- complex with a second humus horizon (Prokashev, 1999; et al.) or with buried profiles;
- alluvial;
- volcanic;
- permafrost;
- gleyed (Ag, G);
- subaqueous (shoals, floodplain silty marshes);
- agro-subaqueous (rice bays, etc.);
- primary emerging micro-organoprofile (Abakumov, 2011; Tomashunas, Abakumov, 2014; Lessovaia et al., 2008);
- naturally disturbed mixed where SOM is present: Aeolian, landslide, buried, pit-and-mound, etc. — with subtypes (Karpachevskij et al., 1984; Vasenev, Targul’yan, 1995; Bobrovskij, 2010);
- technogenic on various substrates where SOM is present (Uzhegova, Mahonina, 1984; Alekseev, 1990; Androhanov et al., 2000; Mahonina, 2004; Reintam et al., 2002).
If necessary, the thickness of the organoprofile should be specified: shallow up to 30 cm, thick over 100 cm (the medium-thick category is not designated). The above draft reflects the authors’ ideas and can serve only as a starting point in creating a detailed systematics of soil organoprofiles, taking into account the existing concepts and classifications.
The creation of the classification category «organoprofile» will be justified in soil taxonomy and its further development will obviously be useful in theoretical work on soil evolution. However, despite the long-term existence of this concept (Grishina, Orlov, 1978), it has not been widely used in soil genetic research, this concept is often being reduced to humus forms (Min’kovskij, Shoba, 1994, 1995; Tomashunas, Abakumov, 2014; Bahmet, 2015).
The stated positions on organopedogenesis can be presented in the form of a graphical model of the processes of pedogenesis (Fig. 2), which shows the place of organopedogenesis in the structural organisation of the soil system according to the authors’ ideas.
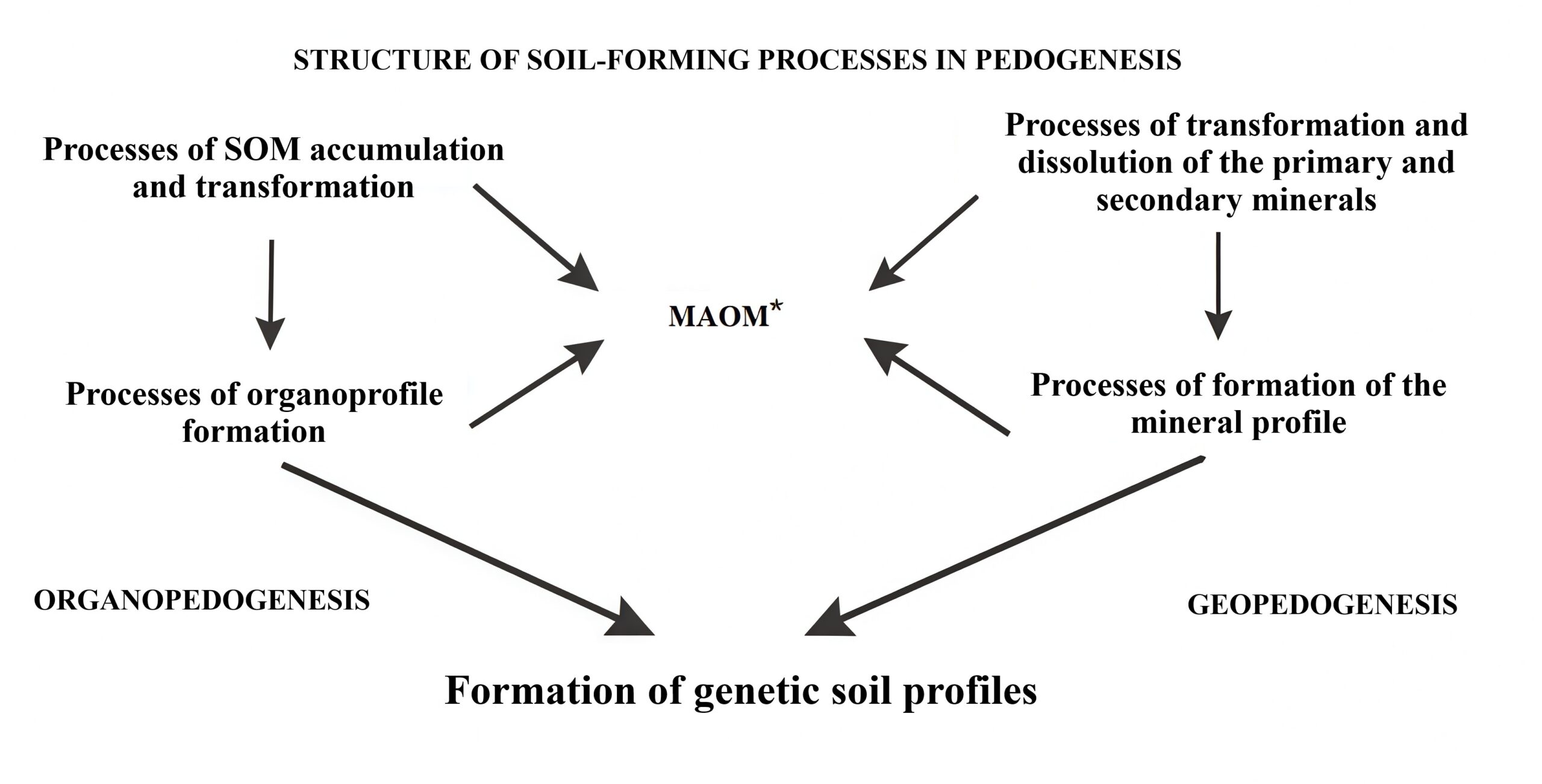
Figure 2. The general structure of pedogenesis at the level of soil formation processes with a reflection of the place of organopedogenesis. The «processes of SOM accumulation and transformation» refer to the humus forms and types of litter; *Mineral associated organic matter including SOM, associated with R2O3 and Ca2+.
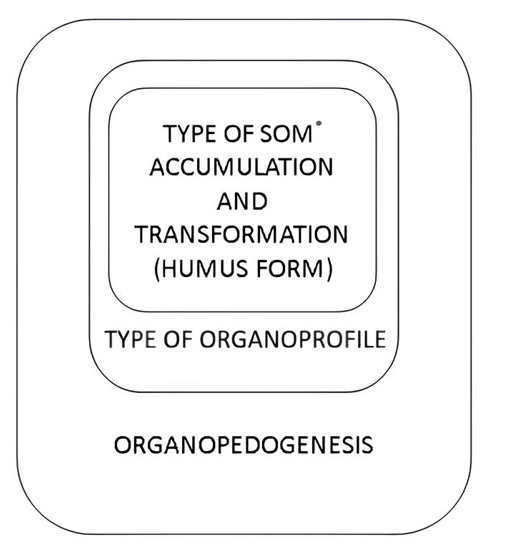
Figure 3. Taxonomic subordination of the components of organopedogenesis («Russian matryoshka doll»); *SOM – soil organic matter
Thus, pedogenesis as a whole encompasses all processes of soil formation, and its hierarchical differentiation into organopedogenesis and geopedogenesis (process affecting only the mineral matrix: the term proposed by the authors) instead of a single soil morphogenesis (Shoba, 1988) seems logical. At the basic hierarchical level, it is logical to identify the categories «Types of organoprofile» and «Types of mineral profile», and below that, «Formation of types of SOM accumulation and transformation», also taking into account the complex processes of SOM migration and the formation of organomineral complexes of different genesis in the soil profile, with release into the biogeochemical cycle at the landscape, regional and global levels.
From the point of view of pedogenesis organisation, another form of presentation, the «Russian matryoshka doll» (Fig. 3) will be useful, allowing us to understand the subordination of the structural components of general pedogenesis, taking into account the role of SOM formation. In this interpretation, the types of litter and other organogenic horizons of automorphic positions logically fit into the general structure of organopedogenesis.
Rates of organopedogenesis
Figure 3 can also be used to demonstrate the rate of formation of organopedogenesis components. The type of SOM accumulation/transformation (humus forms) is a dynamic structure that includes the most rapidly forming subhorizons of the plant litter, the OL litter subhorizon and the OF fermentation subhorizon with formation rates from 1 to 100 years, and the OH humified subhorizon of the litter plus the Ah horizon with morphogenesis times from 10 to 300 years. These edaphic components responsible for the biotic cycle of C, N, P and other nutrients. They have been studied in detail separately (Min’kovskij, Shoba, 1994; Bogatyryov et al., 2004; Lukina, Nikonov, 1996, 1998; Lukina et al., 2019; Solodovnikov, 2025). It is the topsoil that in the first place determines the effective realised soil fertility in natural, especially forest, ecosystems (Chertov, 1981, 1985; Lukina et al., 2019). At the same time, it should be emphasised that in the soils of boreal forests prevailing in Russia, the forest litter is usually quite thick (more than 10 cm unless there have been recent fires) and represented by all subhorizons. The reason for this is the low litter processing rate due to the low quality of the litter (a high C/N ratio) and the low level of biodiversity, small number and biomass of soil saprophagous organisms. The Ah horizon in these soils is by no means always pronounced, especially on rocky sandy moraines. In coniferous-deciduous and broad-leaved forests, on the contrary, the thickness of the litter is low (1–2 cm), the subhorizons of the litter are very indistinct due to active mixing and processing by soil saprophagous organizms. A high-quality litter subhorizon with a low C/N ratio can be seen in autumn if it is not completely processed immediately; at the same time, the Ah horizon is expressed and the transition layer between OH and Ah is also identified.
Thus, the quality of the litter fall plays an essential role in the formation of the types of SOM accumulation and transformation (humus forms). L. G. Bogatyryov and T. V. Fomina (1991) conditionally accepted the litter subhorizon (O1) as the «parent rock» when they calculating the eluvial-accumulative coefficients according to A. A. Rode (1984). In our opinion, the litter can indeed be considered as a second parent rock, since the quality of the litter, as well as the composition of the soil-forming rock, plays a major role in soil formation.
Unlike humus forms, organoprofile is already a soil-genetic category with formation times from 100 years to millennia, playing a major role in the development of a complete soil profile with the transformation and dissolution of minerals due to the energy of SOM mineralized by biota and with the release of organic and mineral substances from biotic cycles into biogeochemical cycles. At the same time, several humus forms can be formed in the same organoprofile. In fact, the humus form is embedded in the organic profile, as the uppermost accumulative part, the starter and driver of the processes of SOM formation. However, combining the humus form with the organoprofile into one category would be conceptually incorrect due to the differences in their role and formation rates. On the other hand, it is quite possible that the types of organoprofiles will serve as the basis for the creation of general types of soil morphogenesis in the context of the ideas of S. A. Shoba (1988). In geopedogenesis, the periods of formation of the complete mineral profile (Targulyan, 2019) are several times longer than the periods of formation of the organoprofile in organopedogenesis.
The rate of restoration of the humus form and organoprofile in case of their physical disturbance (for example, during windstorms) is quite high under the canopy of vegetation, depending on the quality and mass of the litter input and the preservation of fragments of topsoil (Karpachevskij et al., 1984; Razumovskij, 1999; Vasenev, 2008). However, it will be slow in case of the initial absence of vegetation on the exposed parent rock (horizon C), as during the primary ecogenetic succession (Razumovskij, 1999; Abakumov, 2001). In a case of technogenic disturbance, the restoration of the humus form and organoprofile depends on the intensity and mode of disturbance (pollution, recreation), as well as on the properties of the technogenic substrate (Alekseev, 1990; Androhanov et al., 2000). The rates of formation and restoration of the humus forms and organoprofiles discussed above demonstrate the influence of «time», the classical soil formation factor, on the processes of organopedogenesis. Moreover, these parameters also allow us to estimate the maximum reaction time of organopedogenesis components to changes in natural conditions, most of all to modern climate change. Currently, more accurate estimates of these processes in the absence of experimental data and time series are possible using the existing soil and ecosystem models (e.g., Komarov et al., 2017; Shanin et al., 2022;).
Inclusion of organopedogenesis components in soil classification
As noted above, as far back as in the first half of the 20th century, I. V. Tyurin and V. V. Ponomaryova (1940) denoted the humus forms by adding an adjective to the word «soil» (for example, mull soil, raw humus soil). N. L. Blagovidov and G. L. Burkov (1959) recommended and used this approach in the classification of forest soils (for example, moder humus podzolic light loamy…). Since then, this pattern of soil naming by the humus forms has been used in soil and forest typological work in Northwestern European Russia (Chertov, 1966, 1981; Chertov, Dyrenkov, 1973; Chertov et al., 1978; Dyrenkov, 1984; Fedorchuk et al., 2005; Chertov, Nadporozhskaya, 2018), and the forms/types of humus have been described in detail even with a dichotomous field guide (Chertov, 1974).
All the stated positions determine the need for a classification reflecting the role of organopedogenesis in soil formation. There is also some positive experience here (Chertov, 1974, 1981) and, moreover, a methodological approach has been proposed to expand the Russian soil classification to include the humus forms as types of SOM accumulation and transformation (Chertov, 2025).
According to this approach, when the components of organopedogenesis are included in the current Russian soil classification (Klassifikaciya i diagnostika …, 2004; Polevoj opredelitel’ …, 2008), the types of organoprofiles can take the place of a vaguely defined soil subtype, and the types of SOM accumulation and transformation can be integrated on lower layer, since the humus forms represent the most dynamic morphological structure of the soil. For this taxonomic unit, it is proposed to use the level of «rod» (genus) already existing in the Russian classification in accordance with the suggestion of M. I. Gerasimova (Gerasimova, 2018; Hitrov, Gerasimova, 2022), which was introduced to reflect both the role of soil-forming rocks and pollution, as well as for temporary and mechanistic process categories. Integration of the types of SOM accumulation and transformation (humus forms) should be obligatory introduced for all soil taxa below the type level (for which humus forms already exist). Such a taxonomic architecture does not violate the general composition of the genetic soil classification and in no way seeks to replace it, therefore does not require any structural changes in the existing classification (not instead of the genetic one, but together with the genetic one).
The result will be a combination of an edaphological SOM classification with a genetic one for the entire soil, taking into account that the humus of the lower part of the soil profile is more stable and reflects both the history of the soil and modern soil processes. In this case, the soil map is transformed from a strict classical portrait into a vibrant dynamic picture that will require periodic updating (in the range of 10–100 years in forest ecosystems) to track organopedogenesis patterns under the influence of changing natural and anthropogenic factors.
The implementation of the proposals on combining the typological structures of organopedogenesis with genetic taxa will require a substantial analysis of the existing taxonomy for an understandable and acceptable integration of these components, as well as a thorough study of the taxonomy of organoprofile types in the development of the draft proposed above. The algorithm of combining is simple; however, there may be some difficulties here. For example, a situation of uncertainty may arise when it is unclear which type of organoprofile or humus form is represented in the soil under study (for example, in takyr or solonchak). In this case, one should simply omit the indication of the organoprofile and the humus form, as the most logical and simple solution, while it is important to keep the SOM description for analysis and conclusion as to whether this is a new organoprofile or a humus form, or it is a variant of the already existing taxa. It should also be borne in mind that in the Russian genetic classification, the genetic names of soils and humus forms coincide on hydromorphic organogenic soils, and, of course, priority should be given to genetic determination.
The proposed concept of organopedogenesis and the procedure for its inclusion in the soil taxonomy may raise various objections. However, this does not change the postulate of the need for a mandatory and sufficiently detailed reflection of SOM morphology in the soil classification, which is important both from the general theoretical biospheric and from the «applied» edaphic positions. So far, no progress has been observed in this area and it has not even been discussed (Lebedeva et al., 2000; Tonkonogov et al., 2005).
Nevertheless, in forest pedology and forestry, humus forms have long been included in methodological guidelines for mapping forest soils, but, unfortunately, they are not yet used in Russia, although they are used in a large number of other countries (Malysheva et al., 2022). More recently, humus forms have also been introduced into the methodological materials of the most important innovative project of national importance «Unified National System for Monitoring Climatically Active Substances» and the National Forest Inventory of Russia.
CONCLUSION
Firstly, our proposed concept of organopedogenesis can expand the theoretical foundation of modern soil science with an emphasis on the crucial role of the biological factor in the functioning of the pedosphere, including in the context of modern climate change. This includes not only the influence of vegetation, but also, to a large extent, of the entire soil biota. Secondly, it is hoped that it will be able to combine the genetic and edaphic branches of the soil science to create an idea of an integrated rapidly evolving soil system, both from a theoretical biospheric perspective and, especially, from the point of view of its practical use in solving problems of optimising environmental management in the context of drastic environmental changes and increasing anthropogenic pressure on the biosphere of the planet.
The authors express their gratitude to the distinguished reviewers for their valuable comments on the content of this work, which significantly improved the presentation of the stated positions.
FINANCING
State funding contract «Biodiversity and ecosystem functions of forests», registration number № 124013000750-1
REFERENCES
Abakumov E. V., Hronologiya ontogeneza pervichnyh pochv: obzor problemy (Chronology of ontogenesis of primary soils: a review of the problem), Vestnik SPbGU, Ser. 3, 2011, Vyp. 3, pp. 114–120.
Abakumov E. V., Soils of mining areas – natural model of soil formation, Proceed. 1st Intern. Conf. on Soils and Archaeology, Szazhalombatta, Hungary, 2001, pp. 163−165.
Alekseev V. A., Lesnye ekosistemy i atmosfernoe zagryaznenie (Forest ecosystems and atmospheric pollution), Leningrad: Nauka 1990, 200 p.
Androhanov V. A., Ovsyannikova S. V., Kurachev V. M., Technozems: properties, regimes, functioning (Technosols: properties, modes, functioning), Novosibirsk: Nauka, 2000, 200 p.
Bahmet O. N., Strukturno-funkcional’naya organizaciya organoprofilej pochv lesnyh ekosistem Severo-Zapada Rossii (Structural and functional organization of soil organoprofiles of forest ecosystems in Northwest Russia), Avtoref. diss. dokt. biol. nauk (Abst. diss. doct. biol. sci.), Petrozavodsk, 2015, 49 p.
Bioraznoobrazie i funkcionirovanie lesnyh ekosistem (Biodiversity and functioning of forest ecosystems), N. V. Lukina (ed.), Moscow: KMK, 2021, 327 p.
Blagovidov N. L., Burkov G. L., Metodicheskie ukazaniya k proizvodstvu pochvennyh issledovanij i harakteristike uslovij mestoobitaniya lesa (Methodical instructions for the production of soil studies and characterization of forest habitat conditions), Leningrad: LTA, 1959. 30 p.
Bobrovskij M. V., Lesnye pochvy evropejskoj Rossii. Bioticheskie i antropogennye faktory formirovaniya (Forest soils of European Russia. Biotic and anthropogenic factors of formation), Moscow: KMK, 2010, 392 p.
Bogatyryov L. G., Demin V. V., Matyshak G. V., Sapozhnikova V. A., O nekotoryh teoreticheskih aspektah issledovaniya lesnyh podstilok (On some theoretical aspects of forest litter research), Lesovedenie, 2004, No 4, pp. 17–29.
Bogatyryov L. G., Fomina T. V., Harakteristika podstilok sosnyakov Prisurskogo lesnogo massiva (Characteristics of pine forest’s litters of the Prisursky forest massif), Vestnik Moskovskogo universiteta, Seriya 17, Pochvovedenie, 91, No 3, pp. 28–39.
Broll G., Brauckmann H.-J., Overesch M., Junge B., Erber C., Milbert, G., Baize D., Nachtergaele F., Topsoil characterization – Recommendations for revision and expansion of the FAO-Draft (1998) with emphasis on humus forms and biological features, Journal of Plant Nutrition and Soil Science. 2006, Vol. 169, No 3, pp. 453–461.
Budyko M. I., Evolyuciya biosfery (Evolution of the biosphere), Leningrad: Gidrometeoizdat, 1984, 488 p.
Chertov O. G. O smyslovoj strukture naimenovanij taksonov pochvennyh klassifikacij (On the semantic structure soil classification taxa names), Voprosy lesnoj nauki, 2025, Vol. 8, No 1, Article 163, DOI: 10.31509/2658-607x-202581-163
Chertov O. G., Dyrenkov S. A., O klassifikacii lesnyh biogeocenozov Karel’skogo pereshejka (On the classification of forest biogeocenoses of the Karelian Isthmus), Trudy Petrozavodskoj lesnoj opytnoj stacii, 1973, Vol. 2, pp. 74–80.
Chertov O. G., Ekologiya lesnyh zemel’. Pochvenno-ekologicheskoe issledovanie lesnyh mestoobitanij (Ecology of forest lands. Soil-ecological study of forest sites), Leningrad: Nauka, 1981, 192 p.
Chertov O. G., Ekotopy tropicheskogo dozhdevogo lesa (Ecotopes of tropical rain forest), Leningrad: Nauka, 1985, 47 p.
Chertov O. G., Filippov G. V., Mel’nickaya G. B., Krupnomasshtabnoe kartirovanie pochv pri lesoustrojstve, Metodicheskie ukazaniya (Large-scale soil mapping in forest inventory. Methodological Instructions), Leningrad: Len. NII lesn. hoz-va, 1978, 52 p.
Chertov O. G., Mel’nickaya G. B., Lamov A. K., Berg I. E., Opyt rabot po lesnoj tipologii na osnove kartirovaniya pochv i mestoobitanij (Experience of work on forest typology based on mapping of soils and sites), Sbornik statej po lesovodstvu, Leningrad: Len. NII lesn. hoz-va, 1974, No 17, pp. 24–47.
Chertov O. G., Nadporozhskaya M. A., Development and application of humus form concept for soil classification, mapping and dynamic modelling in Russia, Applied Soil Ecology, 2017, Vol. 122, pp. 420–423, DOI: 10.1016/j.apsoil.2017.04.006
Chertov O. G., Nadporozhskaya M. A. Formy gumusa lesnyh pochv: koncepcii i klassifikacii (Humus forms of forest soils: concepts and classifications), Pochvovedenie, 2018, No 10, pp. 1202–1214.
Chertov O. G., Nadporozhskaya M. A., Palyonova M. M., Priputina I. V., Edafologiya v strukture pochvovedeniya i ekosistemnoj ekologii (Edaphology in the structure of soil science and ecosystem ecology), Russian Journal of Ecosystem Ecology, 2018, Vol. 3, No 3, DOI: 10.21685/2500-0578-2018-3-2
Chertov O. G., O ekologicheskih funkciyah i evolyucii pochv (On ecological functions and evolution of soils), Vestnik LGU, Ser. 3, 1990, No 2(10), pp. 75–81.
Chertov O. G., Opredelenie tipov gumusa lesnyh pochv (Determination of humus types in forest soils), Metodicheskie ukazaniya, Leningrad: Len. NII lesn. hoz-va, 1974, 16 p.
Chertov O. G., Razumovskij S. M., Ob ekologicheskoj napravlennosti processov razvitiya pochv (On the ecological orientation of soil development processes), Zhurnal obshchej biologii, 1980, Vol. 62, No 3, pp. 386–396.
Chertov O., Gorbushina A., Deventer B., A model for microcolonial fungi growth on rock surfaces, Ecological Modelling, 2004, Vol. 177, No 3-4, pp. 415–426, DOI: 10.1016/j.ecolmodel.2004.02.011
Chukov S. N., Primenenie metoda yadernogo magnitnogo rezonansa k izucheniyu organicheskogo veshchestva pochv (Application of the nuclear magnetic resonance method to the study of soil organic matter), Vestnik Sankt-Peterburgskogo universiteta. Seriya biologicheskaya, 1997, No 4, pp. 50–59.
Danilov D. A., Yakovlev A. A., Zajcev D. A., Ivanov A. A., Izmeneniya v postagrogennyh pochvah v hode vosstanovleniya drevesnoj rastitel’nosti v usloviyah yugo-zapada Leningradskoj oblasti (Changes in postagrogenic soils during the restoration of woody vegetation in the southwest of the Leningrad region), 2024, Izvestiya SPbLTA, No 251, DOI: 10.21266/2079-4304.2024.251.97-122
Dobrovol’skij G. V., Karpachevskij L. O., Kriksunov E. A., Geosfery i pedosfera (Geospheres and pedosphere), Moscow, GEOS, 2010, 190 p.
Dobrovol’skij G. V., Pedosfera – obolochka zhizni planety Zemlya (The pedosphere is the shell of life on planet Earth), Biosfera, 2009, Vol. 1, No 1, pp. 6–14.
Dokuchaev V. V., Russkij chernozyom: Otchyot Vol’nomu ekonomicheskomu obshchestvu (Russian chernozem: Report to the Free Economic Society), St. Petersburg: Tip. Deklerona i Evdokimova, 1883, 376 p.
Duchaufour Ph., Precis de Pedologie, 2nd edition, Paris: Masson, 1965, 482 p.
Dyrenkov S. A., Struktura i dinamika taezhnyh el’nikov (Structure and dynamics of taiga spruce forests), Leningrad: Nauka, 1984, 176 p.
Duchaufour F., Osnovy pochvovedeniya. Evolyuciya pochv (Fundamentals of Soil Science. Soil Evolution). Moscow: Progress, 1979, 592 p.
Emmer I. M., Humus form and soil development during a primary succession of monoculture Pinus sylvestris forests on poor sandy substrates, Amsterdam, 1995, 135 p.
Evolyuciya pochv i pochvennogo pokrova. Teoriya, raznoobrazie prirodnoj evolyucii i antropogennyh transformacij pochv (Evolution of soils and soil cover. Theory, diversity of natural evolution and anthropogenic transformations of soils), V. N. Kudeyarov, I. V. Ivanov (eds.), Moscow: GEOS, 2015, 928 p.
Evolyuciya, funkcionirovanie i ekologicheskaya rol’ pochv kak komponentov biosfery (Evolution, functioning and ecological role of soils as components of the biosphere), A. O. Alekseev (ed.), Moscow: KMK, 2020, 290 p.
Fedorchuk V. N., Neshataev V. Yu., Kuznecova M. L., Lesnye ekosistemy severo-zapadnyh rajonov Rossii. Tipologiya, dinamika, hozyajstvennye osobennosti (Forest ecosystems of northwestern Russia. Typology, dynamics, and economic characteristics), O. G. Chertov (ed.), St. Petersburg: St. Petersburg Research Institute of Forestry, 2005, 381 p.
Fokin A. D., Dinamicheskaya harakteristika gumusovogo profilya podzolistoj pochvy (Dynamic characteristics of the humus profile of podzolic soil), Izvestiya Timiryazevskoj sel’skohozyajstvennoj akademii, 1975, No 4, pp. 80–88.
Fridland V. M., Pochvy i kory vyvetrivaniya vlazhnyh tropikov (Soils and weathering crusts of the humid tropics), AN SSSR, Pochvennyj in-t im. V. V. Dokuchaeva, Moscow: Nauka, 1964, 312 p.
Gerasimova M. I., Nuzhen li uroven’ roda v klassifikacii pochv Rossii? (Is level ROD (genus) nessesary in Russian soil classification?) Byulleten’ Pochvennogo instituta im. V. V. Dokuchaeva, 2018, Vyp. 95, pp. 90–98, DOI: 10.19047/0136-1694-2018-95-90-98
Glazovskaya M. A., Geohimiya prirodnyh i tekhnogennyh landshaftov SSSR (Geochemistry of natural and technogenic landscapes of the USSR), Moscow: Vysshaya shkola, 1988. 327 p.
Gorshkov V. G., Fizicheskie i biologicheskie osnovy ustojchivosti zhizni (Physical and biological foundations of life sustainability), Moscow, 1995, 470 p.
Green R. N., Trowbridge R. L., Klinka K., Towards a taxonomic classification of humus forms, Forest Science, 1993, Vol. 39, No 1, pp. 1–56.
Grigor’ev G. I., Diagnosticheskie pokazateli stepeni okul’turennosti dernovo-podzolistyh, svetlo-seryh i seryh lesnyh suglinistyh pochv (Diagnostic indicators of the degree of agronomic improvement of sod-podzolic, light gray and gray forest loamy soils), Teoreticheskie osnovy i metody opredeleniya optimal’nyh parametrovsvojstv pochv, Moscow, 1980, pp. 21–29.
Grishina L. A., Gumusoobrazovanie i gumusnoe sostoyanie pochv (Humus formation and humus state of soils), Moscow: MGU, 1986, 244 p.
Grishina L. A., Orlov D. S., Sistema pokazatelej gumusnogo sostoyaniya pochv (System of indicators of humus status of soils), Problemy pochvovedeniya (Sovetskie pochvovedy k II Mezhdunarodnomu kongressu pochvovedov v Kanade, 1978), Moscow: 1978, pp. 42–47.
Hitrov N. B., Gerasimova M. I., Predlagaemye izmeneniya v klassifikacii pochv Rossii: diagnosticheskie priznaki i pochvooobrazuyushchie porody (Proposed changes in the classification of soils of Russia: diagnostic features and parent rocks), Pochvovedenie, 2022, No 1, pp. 3–14, DOI: 10.31857/S0032180X22010087
Humusica – Terrestrial Natural Humipedons, A. Zanella, J. Ascher-Jenull (eds.), Applied Soil Ecology, 2018, Vol. 122, Part I, II, III.
IUSS Working Group WRB. World Reference Base for Soil Resources. International soil classification system for naming soils and creating legends for soil maps, 4th edition, International Union of Soil Sciences (IUSS), Vienna, Austria, 2022, DOI: 10.29003/m4174.978-5-317-07235-3.
Kalinina O., Chertov O., Nadporozhskaya M., Giani L., Properties of soil organic matter of Plaggic Anthrosols from Northwest Germany, Northwest and North Russia, Archives of Agronomy and Soil Science, 2009, Vol. 55, No 5, pp. 477–492, DOI: 10.1080/03650340802637733
Karpachevskij L. O., Les i lesnye pochvy (Forest and forest soils), Moscow: Lesnaya promyshlennost’, 1981, 264 p.
Karpachevskij L. O., Stroganova M. P., Targul’yan V. O., Vasenev I. I., Goncharuk N. Yu., Pochvennye mikrosukcessii v klimaksnyh el’nikah yuzhnoj tajgi (Soil microsuccessions in climax spruce forests of the southern taiga, Istoriya razvitiya pochv SSSR v golocene (History of soil development in the USSR in the Holocene), Pushchino: AN SSSR, 1984, pp. 68–69.
Klassifikaciya i diagnostika pochv Rossii (Classification and diagnostics of soils in Russia), L. L. Shishov, V. D. Tonkonogov, I. I. Lebedeva, M. I. Gerasimova (compl.), Smolensk: Ojkumena, 2004, 342 p.
Klinka K., Krestov P., Fons J., Chourmouzis C., Towards a taxonomic classification of humus forms: Third approximation, Scientia Silvia, 1997, No 9, pp. 1–5.
Kõlli R., Köster T., Interrelationships of humus cover (pro humus form) with soil cover and plant cover: humus form as transitional space between soil and plant, Applied Soil Ecology, 2018, Vol. 123, pp. 451–454.
Komarov A., Chertov O., Bykhovets S., Shaw C., Nadporozhskaya M., Frolov P., Shashkov M., Shanin V., Grabarnik P., Priputina I., Zubkova E., Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing, Ecological Modelling, 2017, Vol. 345, pp. 113–124, DOI: 10.1016/j.ecolmodel.2016.08.007
Kostychev P. A., Pochvy chernozemnoj oblasti Rossii, ih proiskhozhdenie, sostav i svojstva (Soils of the chernozem region of Russia, their origin, composition and properties), St. Petersburg: Izd. Devriena, 1886, 231 p.
Kovda V. A., Biogeohimiya pochvennogo pokrova (Biogeochemistry of soil cover), Moscow, 1985, 493 p.
Kruedener A. A., Osnovy klassifikacii tipov nasazhdenij. Materialy po izucheniyu russkogo lesa (Basics of classification of stand types. Materials on the study of the Russian forest), Petrograd, 1916, 190 p.
Kubiena W. L., The soils of Europe. London: Thomas Murby Co., 1953, 318 p.
Lebedeva H. H., Tonkonogov V. D., Gerasimova M. I., Opyt razrabotki faktornoj klassifikacii pochv (Experience in developing a factor classification of soils), Pochvovedenie, 2000, No 2, pp. 148–157.
Lehmann J., Kleber M., The Contentious Nature of Soil Organic Matter, Nature, 2015, Vol. 528, pp. 60–68, DOI: 10.1038/nature16069
Lessovaia S., Chertov O., Goryachkin S., Specificity of pedogenesis in shallow soils on massive rocks of East Fennoscandia, Agricultural Sciences/Zemès ukio Mokslai, 2008, Vol. 12, No 3, pp. 80–86.
Lukina N. V., Bartalev S. A., Geras’kina A. P., Plotnikova A. S., Gornov A. V., Ershov D. V., … & Ruchinskaya E. V., Rol’ starovozrastnyh lesov v akkumulyacii i hranenii ugleroda (The role of old-growth forests in carbon accumulation and storage), Izvestiya Rossijskoj akademii nauk. Seriya geograficheskaya, 2023, Vol. 87, No 4, рр. 536–557.
Lukina N. V., Geraskina A. P., Kuznecova A. I., Smirnov V. E., Gornov A. V., Shevchenko N. E., Tihonova E. V., Funkcional’naya klassifikaciya lesov: aktual’nost’ i podhody k razrabotke (Functional classification of forests: relevance and approaches to development), Lesovedenie, 2021, No 6, pp. 566–580.
Lukina N. V., Kuznecova A. I., Geraskina A. P., Smirnov V. E., Ivanova V. N., Teben’kova D. N., Gornov A. V., Shevchenko N. E., Tihonova E. V., Neuchtennye faktory, opredelyayushchie zapasy ugleroda v lesnyh pochvah, (Unaccounted factors determining carbon reserves in forest soils), Meteorologiya i gidrologiya, 2022, No 10, pp. 92–110.
Lukina N. V., Nikonov V. V., Biogeohimicheskie cikly v lesah Severa v usloviyah aerotekhnogennogo zagryazneniya (Biogeochemical cycles in northern forests under conditions of aerotechnogenic pollution), Apatity: Kol’skij nauchnyj centr RAN, 1996, 192 р.
Lukina N. V., Nikonov V. V., Pitatel’nyj rezhim lesov severnoj tajgi: prirodnye i tekhnogennye aspekty (Nutrient regime of northern taiga forests: natural and technogenic aspects), Apatity: Kol’skij nauchnyj centr RAN, 1998, 315 р.
Lukina N. V., Orlova M. A., Bahmet O. N., Tihonova E. V., Teben’kova D. N., Kazakova A. I., Kryshen’ A. M., Gornov A. V., Smirnov V. E., Shashkov M. P., Ershov V. V., Knyazeva S. V., Vliyanie rastitel’nosti na harakteristiki lesnyh pochv Respubliki Kareliya (Influence of vegetation on the characteristics of forest soils of the Republic of Karelia), Pochvovedenie, 2019, No 7, pp. 827–842.
Lukina N. V., Orlova M. A., Isaeva L. G., Plodorodie lesnyh pochv kak osnova vzaimosvyazi pochva-rastitel’nost’ (Fertility of forest soils as a basis for soil-vegetation relationships), Lesovedenie, 2010, No 5, pp. 45–56.
Lukina N. V., Polyanskaya L. M., Orlova M. A., Pitatel’nyj rezhim pochv severotaezhnyh lesov (Nutrient regime of soils of northern taiga forests), Moscow: Nauka, 2008, 341 р.
Lukina N. V., Tikhonova E. V., Danilova M. A., Bakhmet O. N., Kryshen A. M., …, & Zukert N. V., Associations between forest vegetation and the fertility of soil organic horizons in northwestern Russia, Forest Ecosystems, 2019, pp. 1–19, doi: 10.1186/s40663-019-0190-2
Lukina N. V., Vliyanie sovremennyh izmenenij klimata na lesnye pochvy (The impact of modern climate change on forest soils), Seminar Instituta global’nogo klimata i ekologii imeni akademika Yu. A. Izraelya, IGKE, 2025, URL: clck.ru/3PJV2o (2025, 01 July).
Mahonina G. I., Nachal’nye processy pochvoobrazovaniya v tekhnogennyh ekosistemah Urala (Initial processes of soil formation in technogenic ecosystems of the Urals), Avtoref. diss. dokt. biol. nauk (Abst. diss. doct. biol. sci.), Tomsk, 2004, 38 p.
Malysheva N. V., Filipchuk A. N., Zolina T. A., Kinigopulo P. S., SHalimova E. M., Popik S. A., Sil’nyagina G. V., Analiz zarubezhnogo opyta nacional’nyh inventarizacij lesov: metody, vyborka, rezul’taty i mezhdunarodnaya statistika (Analysis of foreign experience of national forest inventories: methods, sampling, results and international statistics), Lesohozyajstvennaya informaciya, 2022, No 2, pp. 90–132, DOI: 10.24419/LHI.2304-3083.2022.2.08
Min’kovskij G. M., Shoba S. A., Morfologiya i klassifikaciya organoprofilej pochv yuzhnoj tajgi (Morphology and classification of soil organoprofiles in the southern taiga), Pochvovedenie, 1994, No 9, pp. 90–101.
Min’kovskij G. M., Shoba S. A., Morfostrukturnye podhody k tipizacii organoprofilej pochv (Morphostructural approaches to the typology of soil organoprofiles), Pochvovedenie, 1995, No 10, pp. 1271–1283.
Mirovaya referativnaya baza pochvennyh resursov. Mezhdunarodnaya sistema pochvennoj klassifikacii dlya diagnostiki pochv i sostavleniya legend pochvennyh kart (World database of soil resources. International system of soil classification for soil diagnostics and compilation of soil map legends: translate from English), M. I. Gerasimova, P. V. Krasil’nikov (eds.), 4th edition, Mezhdunarodnyj soyuz nauk o pochve, Moscow: MAKS Press, 2024, 248 p.
Müller P. E., Studien über die natürlichen Humusformen und deren Entwicklung auf Vegetation und Boden, Berlin, 1887, 324 p.
Nadporozhskaya M. A., Mohren G. M. J., Chertov O. G., Komarov A. S., Mikhailov A. V., Dynamics of soil organic matter in primary and secondary forest succession on sandy soils in The Netherlands: An application of the ROMUL model, Ecological Modelling, 2006, Vol. 190, pp. 399–418, DOI: 10.1016/j.ecolmodel.2005.03.025
Odum Yu., Ekologiya (Ecology), Vol. 2, Moscow, Mir, 1986, 376 p.
Orlov D. S., Biryukova O. N., Suhanova N. I., Organicheskoe veshchestvo pochv Rossijskoj Federacii (Organic matter of soils of the Russian Federation), Moscow: Nauka, 1996, 256 p.
Perel’man A. I., Geohimiya landshafta (Geochemistry of the landscape), Moscow: Vysshaya shkola, 1975, 341 p.
Piccolo A., Spaccini R., Savy D., Drosos M., Cozzolino V., The Soil Humeome: Chemical structure, functions and technological perspectives, In: Sustainable Agrochemistry / Vaz Jr. Silvio (ed.), Springer, Cham, 2019, DOI: 10.1007/978-3-030-17891-8_7
Polevoj opredelitel’ pochv (Field soils guide), Moscow: Pochvennyj in-t im. V. V. Dokuchaeva, 2008, 182 p.
Ponge J.-F., Humus forms in terrestrial ecosystems: a framework to biodiversity, Soil Biology and Biochemistry, 2003, Vol. 35, No 7, pp. 935–945, DOI: 10.1016/s0038-0717(03)00149-4.hal-00498465
Ponomareva V. V., Teoriya podzoloobrazovatel’nogo processa (Theory of the podzol-forming process), Moscow-Leningrad: Nauka, 1964, 379 р.
Prokashev A. M., Pochvy so slozhnym organoprofilem yuga Kirovskoj oblasti (Soils with a complex organoprofile in the south of the Kirov region), Kirov, 1999, 176 p.
Razumovskij S. M., Izbrannye Trudy (Selected works), Moscow: KMK, 1999, 560 р.
Reintam L., Elmar K., Rooma I., Development of soil organic matter under pine on quarry detritus of open-cast oil-shale mining, Forest Ecology and Management, 2002, Vol. 171, pp. 191−198.
Rode A. A., Genezis pochv i sovremennye faktory pochvoobrazovaniya (Soil genesis and modern factors of soil formation), Moscow: Nauka, 1984, 256 p.
Rodin L. E., Bazilevich N. I., Dinamika organicheskogo veshchestva i biologicheskij krugovorot zol’nyh elementov i azota v osnovnyh tipah rastitel’nosti zemnogo shara (Dynamics of organic matter and biological cycle of ash elements and nitrogen in the main vegetation types of the globe), Akad. nauk SSSR. Botan. in-t im. V. L. Komarova, Moscow-Leningrad: Nauka, 1965, 253 p.
Semenov V. M, Kogut B. M., Pochvennoe organicheskoe veshchestvo (Soil Organic Matter), Moscow: GEOS, 2015, 233 p.
Semenov V. M., Lebedeva T. N., Sokolov D. A., Zinyakova N. B., Lopes de Gerenyu V. O., Semenov M. V., Izmerenie pochvennyh pulov organicheskogo ugleroda, vydelennyh bio-fiziko-himicheskimi sposobami frakcionirovaniya (Measurement of the soil organic carbon pools isolated using bio-physical-chemical fractionation methods), Pochvovedenie, 2023, No 9, pp. 1155–1172, DOI: 10.31857/S0032180X23600427
Shanin V. N., Frolov P. V., Priputina I. V., Chertov O. G., Byhovec S. S., Zubkova E. V., Portnov A. M., Frolova G. G., Stamenov M. N., Grabarnik P. Ya., Modelirovanie dinamiki lesnyh ekosistem s uchyotom ih strukturnoj neodnorodnosti na raznyh funkcional’nyh i prostranstvennyh urovnyah (Modeling the dynamics of forest ecosystems taking into account their structural heterogeneity at different functional and spatial levels), Voprosy lesnoj nauki, 2022, Vol. 6, No 3, DOI: 10.31509/2658-607x-202252-112
Shoba S. A., Morfogenez pochv lesnoj zony (Morphogenesis of soils in the forest zone), Avtoref. diss. dokt. biol. nauk (Abst. diss. doct. biol. sci.), Moscow, 1988, 48 p.
Shumakov B. C., Principy klassifikacii, nomenklatury i kartirovaniya lesnyh podstilok (Principles of classification, nomenclature and mapping of forest litter), Sbornik rabot po lesnomu hozyajstvu, Vyp. 35, Moscow–Leningrad: Goslesbumizdat, 1958, рр. 199–216.
Sibircev N. M., Pochvovedenie (Soil Science), St. Petersburg: Izd. Skvorcova, 1899, 360 p.
Smirnova O. V., Populyacionnaya organizaciya biocenoticheskogo pokrova lesnyh landshaftov (Population organization of the biocenotic cover of forest landscapes), Uspekhi sovremennoj biologii, 1998, Vol. 118, No 2, pp. 148–165.
Smirnova O. V., Toropova N. A., Potencial’naya rastitel’nost’ i potencial’nyj ekosistemnyj pokrov (Potential vegetation and potential ecosystem cover), Uspekhi sovremennoj biologii, 2016, Vol. 136, No 2, pp. 199–211.
Smirnova O. V., Toropova N. A., Sukcessiya i klimaks kak ekosistemnyj process (Succession and climax as an ecosystem process), Uspekhi sovremennoj biologii, 2008, Vol. 128, No 2, pp. 129–144.
Sokolov I. A., Pochvoobrazovanie i ekzogenez (Soil formation and exogenesis), Moscow, 1997, 244 p.
Solodovnikov A. N., Modeli nachal’nogo razlozheniya hvoi sosny obyknovennoj v srednetaezhnyh usloviyah Vostochnoj Fennoskandii (Models of initial decomposition of Scots pine needles in the middle taiga conditions of Eastern Fennoscandia), Byulleten’ Pochvennogo instituta im. V. V. Dokuchaeva, 2025, Vyp, 122, pp. 41–61, DOI: 10.19047/0136-1694-2025-122-41-61
Sukachev V. N., Dylis N. V., Osnovy lesnoj biogeocenologii (Fundamentals of forest biogeocenology), Moscow: Nauka, 1964, 573 p.
Targul’yan V. O., Teoriya pedogeneza i evolyucii pochv (Theory of pedogenesis and soil evolution), Moscow: GEOS, 2019, 295 p.
Tomashunas V. M., Abakumov E. V., Organoprofili pochv poluostrovov Yamal i Gydan (Soil organoprofiles of the Yamal and Gydan peninsulas), Mezhdunarodnaya nauchn. konf. «Ekologiya i biologiya pochv» (International Scientific Conference «Ecology and Biology of Soils»), Rostov-na-Donu, 2014, pp. 364–365.
Trofimov S. S., Taranov S. A., Naplekova N. N., Fatkulin F. A., Gumusoobrazovanie v tekhnogennyh ekosistemah (Humus formation in pollution-induced ecosystems), Novosibirsk: Nauka, 1986, 126 p.
Tyurin I. V., Ponomaryova V. V., Materialy po izucheniyu gumusa lesnyh pochv (Materials to the forest soils humus investigation), Trudy Leningradskoj lesotekhnicheskoj akademii, 1940, Vyp. 56, p. 3–49.
Uzhegova I. A., Mahonina G. I., Nachal’nye processy pochvoobrazovaniya na otvalah Pervoural’skogo mestorozhdeniya zheleznyh rud (Initial processes of soil formation on the dumps of the Pervouralsk iron ore deposit), Pochvovedenie, 1984, No 11, pp. 14−21.
Vasenev I. I., Pochvennye sukcessii (Soil successions), Moscow: LKI, 2008, 400 p.
Vasenev I. P., Targul’yan V. O., Vetroval i taezhnoe pochvoobrazovanie (Windfall and taiga soil formation), Moscow: Nauka, 1995, 247 p.
Vernadsky V. I., Biosfera (Biosphere) Leningrad: Nauch. him.-tekhnol. izd-vo, 1926, 147 p.
Vernadsky V. I., Zhivoe veshchestvo i biosfera (Living substance and biosphere), Moscow: Nauka, 1994, 671 p.
Wachendorf C., Milbert G., Broll G., Frank T., Graefe U., Beylich A., A concept for a consolidated humus form description – An updated version of the German humus form systematics, International Journal of Plant Biology, 2023, Vol. 14, No 3, pp. 658–686.
Waksman S. A., Gumus: Proiskhozhdenie, himicheskij sostav i znachenie ego v prirode (Humus: Origin, chemical composition and its significance in nature), Moscow: Sel’hozgiz, 1937, 471 p.
Wilde S. A., Forest soils: origin, properties, relation to vegetation and silvicultural management, New York: Wiley & Sons, 1958, 537 p.
Williams V. R., Pochvovedenie. Izbrannye sochineniya (Soil science. Selected works), Moscow: Yurajt, 2025, 344 p.
Yarilov A. A., Pedologiya kak samostoyatel’naya estestvennonauchnaya disciplina o zemle
(Pedology as an independent natural science discipline about the earth), Yur’ev (Tartu): Izd. Yur’evskogo un-ta. 1904 –1905, Part 1, 480 p.; Part 2, 244 p.
Zanella A., Ponge J.-F., Briones M. J. I., Terrestrial humus systems and forms – Biological activity, space-time dynamics, Applied Soil Ecology, 2018, Vol. 122, Part 1, pp. 103–137, DOI: 10.1016/j.apsoil.2017.07.020
Zanella A., Ponge J.-F., Jabiol B., Van Delft B., De Waal R., Katzensteiner K., … & Hager H., Standardized Morpho-Functional Classification of the Planet’s Humipedons, Soil Systems, 2022, Vol. 6, No 3, Article 59, DOI: 10.3390/soilsystems6030059
Reviewers: D. Sc. (Biology), Corresponding Member, Russian Academy of Sciences (RAS) P. S. Krasilnikov, D. Sc. (Biology), Professor M. I. Gerasimova
[1] This paper is based on the results of many years of research by the authors, therefore it refers to a significant number of their publications, as well as contains references to long-standing works of classics, which is necessary to highlight the proposed approach. The authors are grateful to the editors and reviewers for their understanding.
[2] The term «organogenesis» is more concise, but it has been already used in zoology and medicine, where it stands for the formation of organs in embryos. By the way, the term «pedogenesis» meaning one of the types of parthenogenesis, has been also used in zoology since the 19th century.





