- DOI 10.31509/2658-607x-2019-2-2-1-49
Classification of forests using a field guide of forest types of the European Russia (evidence from Karelia and the Karelian isthmus)
С текстом оригинальной статьи Вы можете ознакомиться по ссылке
![]() © 2019 г.
© 2019 г.
A.V. Gornov
Center for Forest Ecology and Productivity of the RAS
Profsoyuznaya st. 84/32 bldg. 14, Moscow, 117997, Russia
E-mail: aleksey-gornov@yandex.ru
Received 15 November 2018
Classification of forests of Karelia and the Karelian Isthmus was carried out using the field guide of the forest types of the European Russia developed by L. B. Zaugolnova and V. B. Martynenko. The studied forests were divided into five main sections: lichenous, green moss, sphagnous, grass and grassmarsh. The most common sections were the green moss and the sphagnous one. They include five groups of forest types each. The least diverse is the lichenous section: it consists of only two groups of forest types. In each group of forest types individual forest types were identified. The most common types of forest were bilberry-green moss pine and spruce forests.
Key words: classification of forests, field guide of forest types of the European Russia, Karelia, Karelian Isthmus, northern taiga, middle taiga
One of the first stages of studying forest vegetation is classifying it. A number of approaches to forest classification has now been developed. They tend to be either dominant or floristic (Braslavskaya, Zaugolnova, 2010). The dominant approach is based on the analysis of similarities/differences of forest communities only by predominant species from different layers taking into account a limited number of species. The floristic approach implies the analysis of similarities/differences between forest communities based on the entire species composition of each layer (Braun-Blanquet, 1964; Alexandrova, 1969; Mirkin et al., 1989; Mirkin, Naumova, 1998; Bulokhov, Solomeshch, 2003). Both approaches have their advantages that make them important for the practical use of classifications. For example, the relative simplicity of the diagnostic rules of the dominant approach makes it useful for solving practical problems. The ecological and floristic classification based on the species association can be used to assess the ecological regime of communities. All of the above encouraged some scientists to develop the ecological-coenotic approach to forest classification (Zaugolnova, Morozova, 2006). As a result, L. B. Zaugolnova and V. B. Martynenko created an electronic field guide of the forest types in the European Russia (http://cepl.rssi.ru/bio/forest/index.htm). This field guide allows describing the typological diversity of the forests, according to the practice of domestic forest phytocenology and foreign approaches to plants classification. This helps to set up correspondences between syntaxons in ecological and floristic classification and forest types in the dominant classification. In this regard, the aim of our study was to classify the forests of Karelia and the Karelian Isthmus using the field guide of the forest types of the European Russia.
Materials and methods
The material was collected in the forests of Karelia and the Karelian Isthmus as part of the ICP Forests international program (Lukina et al., 2013). The study area is fully located in the taiga zone. Karelia is divided into two unequal parts – the middle and northern subzones of the taiga. In the middle taiga of Karelia 42 permanent lookout points (PLPs) were established, and in the northern taiga – 58 lookout points (Fig. 1). The Karelian Isthmus was subject to various geobotanical zonings: it belonged entirely to the southern taiga (Yurkovskaya, Elina, 2009) or was divided into southern and middle subzones (Geobotanical…, 1989). Despite the fact that the territory of the Karelian Isthmus is much smaller in area than the middle and northern taiga parts of Karelia, 47 PLPs were established there. The geographical location of the territory to a large extent explains the species and cenotic diversity of forest ecosystems, as well as their dynamics; diversity indicators of are considered separately for the Karelian Isthmus, the middle and northern subzones of the Karelian taiga.
The ground cover study was performed by the forest monitoring method according to the ICP Forests international program (2008). 4 plots of 100 sq. m. each were randomly made within the PLPs. The plots were usually square in shape. If due to local conditions it was impossible to establish a square-shaped site, it was made rectangular. The estimation plots were located as far as possible from each other within the boundaries of the PLP or its buffer zone (Fig. 2) to ensure that the long-term similarity (if any) between the estimation plots occurs not only due to spatial coincidences. Geobotanical descriptions are made for 147 PLPs.
Geobotanical descriptions were made by T. Yu. Braslavskaya (Center for Forest Ecology and Productivity of the RAS), N. V. Genikova (Forest Institute of the Karelian scientific centre of the RAS), E. E. Kostina (Forest Institute of the Karelian scientific center of the RAS) and N.V. Ivanova. Geobotanical description of plots for estimation included a complete floristic list taking into account the layer structure of the vegetation. The moss and lichen layer (D) includes ground-storey bryophytes and lichens; the grass and dwarf shrub layer (C) includes all herbaceous and woody plants of over 0.5 m high (dwarf shrubs, dwarf semishrubs, seedlings and young trees); the layer of shrubs and understorey of trees (B) includes shrubs and young trees of more than 0.5 but less than 5 m high, and all vines climbing up to this height; the tree layer (A) includes woody plants higher than 5 m and all the vines climbing up to this height. In the tree layer as well as in the layer of shrubs and understorey of trees, crown density of all participating species was determined, and in the grass-dwarf shrub and moss-lichen layers the projective cover was found and recorded as the whole number of percent. The main requirement to the description of an estimation plot was the fullest possible identification of the species composition of the vegetation present. Latin names of vascular plants are given according to S. K. Cherepanov (1995).
Classification of vegetation of PLPs. Based on the descriptions of the plots for estimation, each PLP was attributed to a specific forest type. The classification was carried out using the electronic field guide of forest types in the European Russia developed by L. B. Zaugolnova and V. B. Martynenko (http://www.cepl.rssi.ru/cepl.htm). In addition, each PLP was provided with a summary geobotanical description containing the information from the four estimation plots.
The field guide covers slightly and moderately disturbed forests with well-developed ground cover (both grass-dwarf shrub and moss-lichen). The principles of both dominant and floristic classifications of the forest communities are taken into account when compiling the field guide. To describe forests within the framework of the dominant approach, the dominance of both individual species (mainly trees) and ecological-coenotic groups of species (for plants of ground layers) was used. A section that unites the communities by the nature of the ground layers is the largest unit in the typological scheme. A name of a section corresponds to the predominant life form of plants in the ground layers. Sections and subsections are named by traditional Russian geobotanical names. A subsection is an association of forest communities within one section, differing by the structure of the grass-dwarf shrub layer. The set of subsections in its most general form reflects the ecological series of zonal complexes of forest vegetation; they can become the basis for the ecological series of individual landscapes and catchment areas to be formed. The main typological unit is a group of forest types. It is a subdivision of subsection communities depending on the predominant species of the tree layer: each group of forest types is characterized by the predominant species of the upper layer of the forest stand combined with a certain ratio of ecological-coenotic groups of species as part of grass-dwarf shrubs and moss-lichen layers. A group of forest types is a set of smaller units, i.e. forest types. A forest type is the forest community the size of which most corresponds to the association as seen in the dominant approach and it is the smallest unit in this typology.
Evaluation of the floristic diversity. For forests species diversity assessment the Whittaker indicators that have become classical in modern ecology: alpha and beta diversity (Mirkin et al., 1989), were used. Geobotanical descriptions made on the estimation plots of the PLPs served as the material for the analysis of floristic diversity.
Alpha diversity is indicative of the richness of species in particular communities. Key indicators of alpha diversity: species richness is the total number of species in a community, and species density is the average number of species per unit area. Simultaneous recording of species richness and species density results in comparable estimates of species diversity in the analysis of different communities such as forest types.
Beta diversity is indicative of the variability of alpha diversity indicators in space during the transition from one type of forest to another. Beta diversity was estimated using the similarity index (Jaccard index) and the heterogeneity index (Whittaker index). In addition to these indices, vegetation beta diversity can be characterized by the set and number of forest types represented in a particular area.
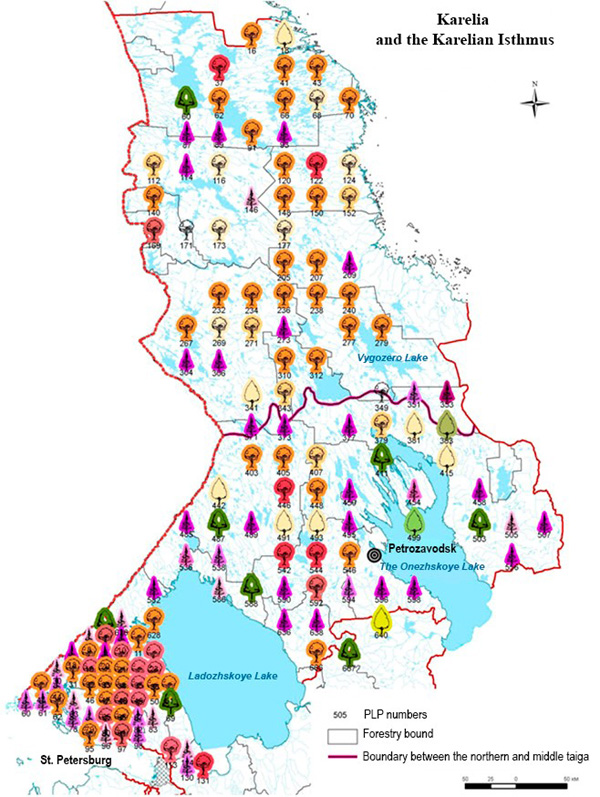
Figure 1. Location of permanent lookout points (PLPs) on the territory of Karelia and the Karelian Isthmus
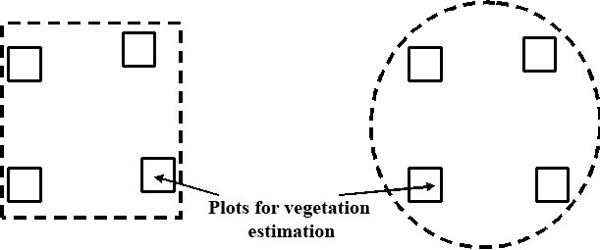
Figure 2. Layout examples of plots for estimation in the territory of PLPs
Ecological-coenotic structure. The structure of species diversity was evaluated by the ratio of species belonging to different ecological-coenotic groups (ECGs). According to A. A. Nitsenko (1969) and G. M. Zozulin (1973), ecological-coenotic groups are large groups of ecologically related species, which are in their origin associated with different types of communities. The study used the classification of ecological-coenotic groups of vascular plant species of the European Russia (Eastern European…, 2004) developed on the basis of ecological-coenotic suites by A. A. Nitsenko and historical suites by G. M. Zozulin. Estimation of the ecological-coenotic structure of communities was carried out according to the general list of species found on all plots for estimation of the PLPs. The generalized version of ECGs was used for the analysis of the forest vegetation: nemoral (Nm), boreal (Br), nitrophilous (Nt), pine forest (Pn), meadow and meadow-marginal (Md), wetland (W).
Results and discussion
According to the geobotanical descriptions of the PLPs, the studied forests of the northern and middle taiga can be divided into 5 main sections depending on the general coverage of the moss-lichen layer and the ratio of lichens, green and sphagnum mosses in its composition: lichenous section (cladinosa), green moss section (hylocomiosa), sphagnous section (sphagnosa), grass section (herbosa), grass and marsh section (uliginosoherbosa). Let us have a more detailed look at them.
Green moss section (hylocomiosa)
Communities of the green moss section are most common in the studied forests: the green moss section comprises 70% of all established PLPs. 40% of these forests are forests of the northern taiga, and 60% are forests of the middle taiga. They are mainly various pine forests. The forests of the green moss section are characterized by a closed tree layer and well-developed moss and lichen cover. As a rule, the projective cover of the moss-lichen layer is at least 50% with green mosses dominating therein. Due to the formation of leaf litter, the moss cover can fall to 40% and less, such communities are a transitional form between the green moss and grass sections. The grass-dwarf shrub layer consists mainly of boreal species: Vaccinium myrtillus, V. vitis-idaea, V. uliginosum, Empetrum hermaphroditum, E. nigrum, Linnaea borealis, etc. At the same time, species of the Empetrum genus are most often found in the forests of the northern taiga.
The green moss section includes forests of five groups of forest types: dwarf shrub-green moss birch forests, small grass-green moss spruce forests, dwarf shrub-green moss spruce forests, dwarf shrub-green moss pine forests and small grass-green moss pine forests (Fig. 3). The most common group of forest types is the group of dwarf shrub-green moss pine forests. They occupy about 50% of the PLPs belonging to the green moss section. The second most common group includes dwarf shrub-green moss spruce forests (about 30% of the PLPs of the green moss section). The share of small grass-green moss spruce and pine forests as well as dwarf shrub-green moss birch forests is approximately the same. Let us consider the groups of forest types of the green moss section.
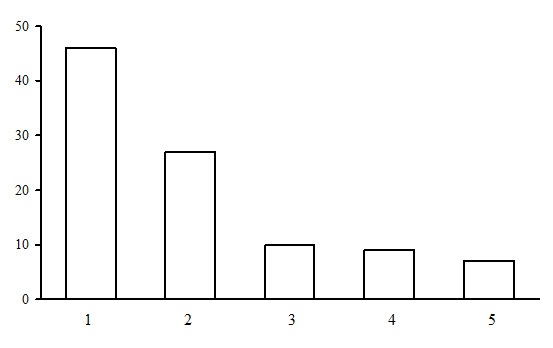
Figure 3. Ratio of PLPs by forest type groups. Green moss section. The X-axis shows groups of forest types (1 – dwarf shrub-green moss pine forests, 2 – dwarf shrub-green moss spruce forests, 3 – small grass-green moss spruce forests, 4 – small grass-green moss pine forests, 5 – dwarf shrub-green moss birch forests), the Y-axis shows the percentage
Dwarf shrub-green moss pine forests are the forests where the tree layer is dominated by Pinus sylvestris (geobotanical description – see Table 1) with Betula pendula, B. pubescens, Picea abies, etc. as common admixtures (Fig. 4).

Figure 4. Dwarf shrub-green moss pine forest
The shrubs layer and the understorey consist of the predominant younger generation of Betula pendula, B. pubescens, Picea abies, Pinus sylvestris trees and accompanying shrub species Juniperus communis, Sorbus aucuparia, Salix caprea, etc. Vaccinium myrtillus and V. vitis-idaea dominate in the grass-dwarf shrub layer both in the northern and middle taiga. Constant species with low coverage: Avenella flexuosa, Linnaea borealis, Lycopodium annotinum, Luzula pilosa, Equisetum sylvaticum, etc. At the same time, dwarf shrub-green moss pine forests of the northern taiga differ from the same forests of the middle taiga in the composition of the grass-dwarf shrub layer. In the northern taiga the coverage and occurrence of some dwarf shrubs are relatively high: Empetrum nigrum, Ledum palustre, and Calluna vulgaris. In the pine forests of the middle taiga there are species that do not occur or are very rare in pine forests of the northern taiga: Convallaria majalis, Dryopteris carthusiana, Fragaria vesca etc.; Calamagrostis arundinacea can be seen rather constantly. In the middle taiga the fern Pteridium aquilinum can also be found which is a marker of frequent ground fires. The moss-lichen layer is well developed. It is dominated by green mosses: Hylocomium splendens, Pleurozium schreberi, species of the genus Dicranum. Polytrichum commune can have low coverage, but is relatively common. The share of lichens in modest.
Within the dwarf shrub-green moss pine forest group of forest types five forest types are identified: bilberry-green moss pine forests, cowberry-green moss pine forests, cowberry-heather-green moss pine forests, bilberry-ledum-green moss pine forests, and heathberry-bilberry-green moss pine forests. Of those, bilberry-green moss pine forests are predominant, with approximately the same share in the northern and middle taiga (Fig. 5). The northern taiga has two types of forests that are not found in the middle taiga: bilberry-ledum-green moss and heathberry-bilberry-green moss pine forests.
Dwarf shrub-green moss pine forests are almost ubiquitous and are very common in the northern and middle taiga. These forests usually result from repetitive fires. If there are no fires for a long time, the forests turn into pine stands with a large share of spruce, and later – into spruce forests with singular pine trees.
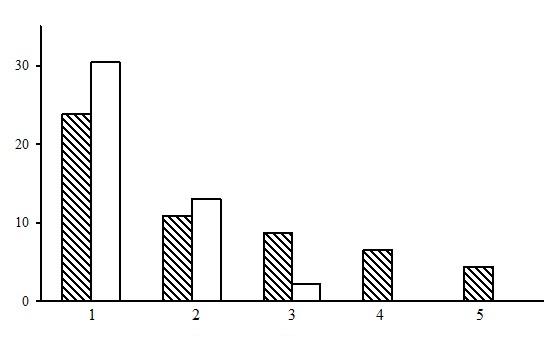
Figure 5. The ratio of forest types in the group of dwarf shrub-green moss pine forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows forest types (1 – bilberry-green moss pine forests, 2 – cowberry-green moss pine forests, 3 – cowberry-heather-green moss pine forests, 4 – bilberry-ledum-green moss pine forests, 5 – heathberry-bilberry-green moss pine forests), the Y-axis shows the percentage
Small grass-green moss pine forests were seen only in PLPs located in the forests of the middle taiga (geobotanical description – see Table 2). In this forest types group, the predominant species of the tree layer is Pinus sylvestris. Betula pendula, B. pubescens and others are usually found as admixtures. Sometimes a second substorey is formed by Picea abies and species of the genus Betula. The layer of shrubs and tree understorey consists of the predominant (Sorbus aucuparia, Frangula alnus, Picea abies) and the accompanying species (Padus avium, Juniperus communis); almost no younger generation of Pinus sylvestris is observed. In the grass-dwarf shrub layer Vaccinium myrtillus, Avenella flexuosa, Oxalis acetosella and others are codominants. In some places, Pteridium aquilinum becomes predominant showing the fire origin of such communities. Calamagrostis arundinacea, Dryopteris carthusiana, Fragaria vesca, Trientalis europaea, and Rubus saxatilis show high occurrence but insignificant abundance. Nemoral species are observed: Convallaria majalis, Lathyrus vernus, and Melica nutans. The moss-lichen layer is poorly developed: its coverage depends on the power of the grass-dwarf shrub layer. Pleurozium schreberi, Hylocomium splendens and species of Dicranum genus are often codominants here.
Table 1. Some geobotanical parameters of the dwarf shrub-green moss pine forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
max |
x |
st. dev. |
|
| Stand age, yrs |
30.0 |
160.0 |
72.6 |
36.5 |
40.0 |
130.0 |
89.3 |
21.5 |
| Stand height, m |
6.0 |
23.0 |
14.4 |
4.0 |
14.0 |
26.0 |
20.7 |
3.1 |
|
Tree canopy density (A), % |
13.8 |
76.3 |
47.3 |
16.1 |
19.3 |
67.5 |
45.0 |
12.1 |
| Density of the layer of shrubs and tree understorey (B), % |
0.0 |
28.8 |
10.0 |
6.8 |
0.1 |
30.0 |
9.7 |
8.4 |
| Total projective cover of the grass layer (C), % |
2.6 |
84.7 |
55.3 |
18.1 |
5.5 |
72.7 |
36.1 |
17.0 |
| Total projective cover of the moss-lichen layer (D), % |
18.8 |
93.1 |
57.9 |
24.0 |
0.6 |
96.6 |
67.7 |
26.2 |
| Total projective cover of green mosses in the moss-lichen layer, % |
18.8 |
80.6 |
51.1 |
20.0 |
0.6 |
96.6 |
65.5 |
25.7 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
27.5 |
5.8 |
8.3 |
0.0 |
13.6 |
1.2 |
3.1 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
10.0 |
1.0 |
2.7 |
0.0 |
17.4 |
1.0 |
3.8 |
|
Number of PLPs |
25 |
21 |
||||||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | ||||||||
Two forest types are subdivided within the group of dwarf shrub-green moss pine forests: bilberry-oxalis-green moss pine forests (8 PLPs) and hair grass-bracken-green moss pine forests (1 PLP).
Small grass-green moss pine forests grow on flat parts and slopes of lake terraces, on the slopes of the dividing ridge on the outwash sandy loam or sandy moraine.
Table 2. Some geobotanical parameters of the small grass-green moss pine forests
|
Parameters |
Forest area |
|||
|
Middle taiga |
||||
|
min |
Max |
x |
st. dev. |
|
| Stand age, yrs |
70.0 |
120.0 |
94.4 |
15.1 |
| Stand height, m |
21.0 |
27.0 |
25.0 |
2.4 |
|
Tree canopy density (A), % |
22.5 |
85.3 |
57.1 |
19.1 |
| Density of the layer of shrubs and tree understorey (B), % |
1.6 |
28.6 |
18.6 |
7.9 |
| Total projective cover of the grass layer (C), % |
15.6 |
81.1 |
59.0 |
20.8 |
| Total projective cover of the moss-lichen layer (D), % |
0.9 |
31.4 |
11.1 |
11.2 |
| Total projective cover of green mosses in the moss-lichen layer, % |
0.9 |
31.3 |
9.7 |
11.3 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
0.5 |
0.1 |
0.2 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
11.8 |
1.4 |
3.9 |
|
Number of PLPs |
9 |
|||
|
Comment. Legend – see Table 1. |
||||

Figure 6. Dwarf shrub-green moss spruce forest
Dwarf shrub-green moss spruce forests (geobotanical description – see Table. 3). Picea abies is dominating in the tree layer with the admixture of Betula pubescens (Fig. 6). The layer of tree understorey and shrubs consists of Picea abies, Betula pubescens, Sorbus aucuparia, Juniperus communis, etc. The grass-dwarf shrub layer of spruce forests of the northern and middle taiga is usually dominated by Vaccinium myrtillus and V. vitis-idaea. Constant species: Avenella flexuosa, Solidago virgaurea, Linnaea borealis, etc. At the same time, the forests of the northern taiga have a relatively high coverage and occurrence of some dwarf shrubs: Empetrum nigrum and Vaccinium uliginosum. The moss-lichen layer is formed by Pleurozium schreberi, Hylocomium splendens, and Dicranum scoparium. Polytrichum commune is a constant species with modest abundance. Microdepressions are home to some species of Sphagnum genus. The share of lichens within this layer is small.
Three types of forests are subdivided within the group of dwarf shrub-green moss spruce forests: bilberry-green moss spruce forests, cowberry-green moss spruce forests and dwarf shrub-green moss spruce forests. Of those, bilberry-green moss spruce forests are predominant, most often to be seen in the middle taiga (Fig. 7).
Table 3. Some geobotanical parameters of the dwarf shrub-green moss spruce forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
Max |
x |
st. dev. |
|
| Stand age, yrs |
30.0 |
190.0 |
101.8 |
52.4 |
40.0 |
230.0 |
100.0 |
41.3 |
| Stand height, m |
7.0 |
21.0 |
14.0 |
4.4 |
11.0 |
26.0 |
19.3 |
3.9 |
|
Tree canopy density (A), % |
15.0 |
71.3 |
38.9 |
18.6 |
0.0 |
80.5 |
44.0 |
30.6 |
| Density of the layer of shrubs and tree understorey (B), % |
2.5 |
67.5 |
14.4 |
19.2 |
0.0 |
15.0 |
5.5 |
3.9 |
| Total projective cover of the grass layer (C), % |
29.8 |
77.2 |
46.3 |
15.5 |
8.1 |
60.8 |
35.8 |
13.9 |
| Total projective cover of the moss-lichen layer (D), % |
56.7 |
91.3 |
76.5 |
12.4 |
12.1 |
94.3 |
54.6 |
24.4 |
| Total projective cover of green mosses in the moss-lichen layer, % |
37.9 |
85.0 |
63.0 |
15.9 |
12.1 |
87.2 |
46.6 |
21.3 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
19.2 |
3.2 |
6.0 |
0.0 |
7.1 |
0.8 |
2.0 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
35.8 |
10.2 |
11.9 |
0.0 |
24.6 |
7.1 |
9.1 |
| Number of PLPs |
11 |
16 |
||||||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | ||||||||
Dwarf shrub-green moss spruce forests are virtually ubiquitous and rather common in the northern and middle taiga. Both spruce and pine forests often have post-fire origin.
Small grass-green moss spruce forests (geobotanical description – see Table 4). In the entire network of PLPs located both in the northern and middle taiga this group of forest types was found on ten plots: one in the forests of the northern taiga and nine – in the middle taiga. The tree layer is formed by Picea abies with a small admixture of Betula pubescens, B. pendula, Pinus sylvestris, and less frequently – Alnus incana (Fig. 8). A mixed tree layer of Picea abies and Pinus sylvestris may occasionally be formed. In this case, the structure and composition of lower layers may resemble similar groups of spruce and pine forests, depending on the advancement of secondary (usually post-fire) successions from pine forests to spruce forests. The layer of shrubs and understorey consists of Picea abies, Sorbus aucuparia, Betula pubescens, B. pendula, Alnus incana, etc. Regeneration of Picea abies is associated with microhills and the periphery of the «windows» in the tree canopy. The grass-dwarf shrub layer is codominated by Oxalis acetosella, Vaccinium myrtillus, Avenella flexuosa, sometimes Gymnocarpium dryopteris; Calamagrostis arundinacea, Maianthemum bifolium, Equisetum sylvaticum, and Trientalis europaea can be found with high constancy. Several nemoral species of low abundance are typically present: Convallaria majalis, Stellaria holostea, Lathyrus vernus, Melica nutans, etc. The share of short grasses in the grass and dwarf shrub layer is higher as compared to dwarf shrub-green moss spruce forests. The degree of development of this layer depends on the closeness of tree crowns and the development of the layer of shrubs and the understorey. The moss and lichen layer is characterized by the codominance of Hylocomium splendens, Pleurozium schreberi, and Dicranum species. Microdepressions are home to some species of Sphagnum genus. The share of lichens in the composition of the layer is insignificant.
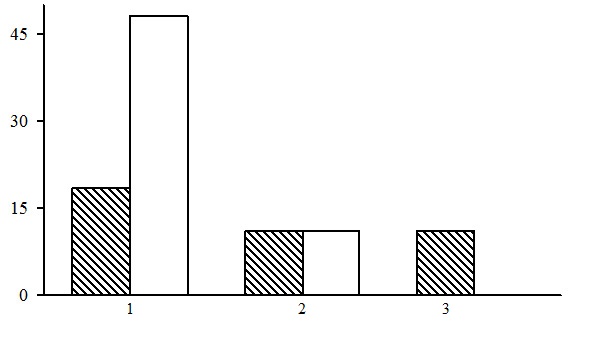
Figure 7. The ratio of forest types in the group of dwarf shrub-green moss spruce forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows forest types (1 – bilberry-green moss spruce forests, 2– cowberry-green moss spruce forests, 3 – dwarf shrub-green moss spruce forests), the Y-axis shows the percentage
Dwarf shrub-green moss spruce forests are virtually ubiquitous and rather common in the northern and middle taiga. Both spruce and pine forests often are of post-fire origin.
Small grass-green moss spruce forests (geobotanical description – see Table 4). In the entire network of PLPs located both in the northern and middle taiga this group of forest types was found on ten plots: one in the forests of the northern taiga and nine – in the middle taiga. The tree layer is formed by Picea abies with a small admixture of Betula pubescens, B. pendula, Pinus sylvestris, and less frequently – Alnus incana (Fig. 8). A mixed tree layer of Picea abies and Pinus sylvestris may occasionally be formed. In this case, the structure and composition of lower layers may resemble similar groups of spruce and pine forests, depending on the advancement of secondary (usually post-fire) successions from pine forests to spruce forests. The shrubs layer and the understorey consist of Picea abies, Sorbus aucuparia, Betula pubescens, B. pendula, Alnus incana, etc. Regeneration of Picea abies is associated with microhills and the periphery of the «windows» in the tree canopy. The grass-dwarf shrub layer is codominated by Oxalis acetosella, Vaccinium myrtillus, Avenella flexuosa, sometimes Gymnocarpium dryopteris; Calamagrostis arundinacea, Maianthemum bifolium, Equisetum sylvaticum, and Trientalis europaea can be found with high constancy. Several nemoral species of low abundance typically present: Convallaria majalis, Stellaria holostea, Lathyrus vernus, Melica nutans, etc. The share of short grasses is higher in the grass and dwarf shrub layer as compared to dwarf shrub-green moss spruce forests. The degree of development of this layer depends on the closeness of tree crowns and the development of the shrubs layer and the understorey. The moss and lichen layer is characterized by the codominance of Hylocomium splendens, Pleurozium schreberi, and Dicranum species. Microdepressions are home to some species of Sphagnum genus. The share of lichens in the composition of the layer is insignificant.

Figure 8. Small grass-green moss spruce forest
Within the group of small grass-green moss spruce forests three types of forests are identified: oxalis-fern spruce forests, spruce bilberry-small grass-green moss spruce forests, and fern-green moss spruce forests. Of these, bilberry-small grass-green moss spruce forests dominate, being common in the middle taiga (Fig. 9).
Table 4. Some geobotanical parameters of the small grass-green moss spruce forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
| Stand age, yrs |
250.0 |
65.0 |
130.0 |
91.7 |
17.3 |
| Stand height, m |
20.0 |
22.0 |
29.0 |
24.8 |
2.9 |
|
Tree canopy density (A), % |
60.0 |
0.0 |
89.5 |
54.8 |
24.3 |
| Density of the layer of shrubs and tree understorey (B), % |
8.8 |
2.0 |
22.5 |
9.4 |
7.0 |
| Total projective cover of the grass layer (C), % |
64.0 |
10.8 |
97.1 |
52.5 |
25.4 |
| Total projective cover of the moss-lichen layer (D), % |
52.3 |
0.1 |
82.7 |
31.6 |
31.4 |
| Total projective cover of green mosses in the moss-lichen layer, % |
52.2 |
0.1 |
82.6 |
29.1 |
29.6 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
0.0 |
0.1 |
0.0 |
0.0 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
0.0 |
15.8 |
2.5 |
5.2 |
|
Number of PLPs |
1 |
9 |
|||
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. |
|||||
Table 5. Some geobotanical parameters of the dwarf shrub-green moss birch forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
max |
x |
st. dev. |
|
| Stand age, yrs |
25.0 |
55.0 |
42.0 |
16.1 |
25.0 |
50.0 |
38.8 |
11.1 |
| Stand height, m |
7.0 |
15.0 |
11.2 |
4.0 |
9.0 |
18.0 |
14.8 |
4.0 |
|
Tree canopy density (A), % |
33.8 |
62.0 |
46.3 |
14.8 |
27.5 |
68.8 |
51.9 |
17.6 |
| Density of the layer of shrubs and tree understorey (B), % |
13.8 |
36.5 |
23.2 |
12.7 |
3.5 |
8.0 |
6.5 |
2.0 |
| Total projective cover of the grass layer (C), % |
40.6 |
60.2 |
51.2 |
10.1 |
23.8 |
40.8 |
32.3 |
7.6 |
| Total projective cover of the moss-lichen layer (D), % |
17.9 |
76.6 |
49.0 |
29.8 |
18.7 |
91.7 |
45.1 |
32.1 |
| Total projective cover of green mosses in the moss-lichen layer, % |
17.3 |
52.3 |
38.2 |
20.0 |
18.3 |
80.2 |
41.9 |
26.8 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
24.3 |
9.7 |
14.0 |
0.0 |
11.5 |
2.9 |
5.7 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
4.3 |
1.8 |
2.3 |
0.0 |
0.8 |
0.3 |
0.4 |
|
Number of PLPs |
3 |
4 |
||||||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | ||||||||
Small grass-green moss spruce forests often grow on drained habitats; they can be found on the slopes of moraine hills, on flat sections of dividing ridges, on the smooth slopes of the foothills on both noncalcareous and calcareous moraine loams. This group of forest types has traditionally been regarded as representing zonal vegetation of the middle taiga; these forests, having higher floristic diversity differ from dwarf shrub-green moss spruce forests.
Dwarf shrub-green moss birch forests (geobotanical description – see Table 5). The forest stand is formed by Betula pubescens with a small admixture of B. pendula, Pinus sylvestris and Picea abies (Fig. 10). The layer of understorey of trees and shrubs consists mainly of Picea abies. Other species (Salix caprea, Pinus sylvestris, Sorbus aucuparia, Juniperus communis, etc.) show low abundance. Vaccinium myrtillus, V. uliginosum, V. vitis-idaea dominate in the grass-dwarf shrub layer both in the northern and middle taiga. Linnaea borealis, Lycopodium annotinum, Trientalis europaea, etc. demonstrate high constancy but insignificant abundance. Sometimes Gymnocarpium dryopteris is present. The presence of Empetrum nigrum is typical for the PLPs located in the northern taiga. The moss-lichen layer is dominated by Pleurozium schreberi, Hylocomium splendens and Polytrichum commune. Microdepressions are home to some species of Sphagnum genus. The share of lichens in the composition of the layer is insignificant.
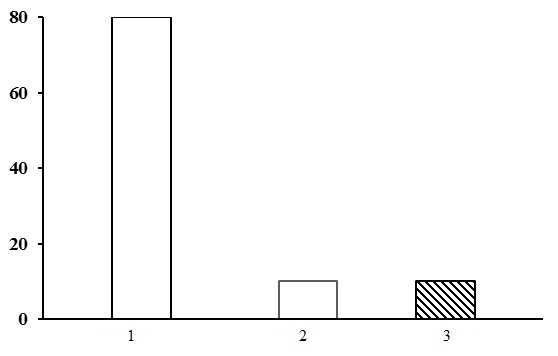
Figure 9. The ratio of forest types in the group of small grass-green moss spruce forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows forest types (1 – bilberry-small grass-green moss spruce forest, 2 – oxalis-fern spruce forest, 3 – small fern-green moss spruce forest),
the Y-axis shows the percentage
Two forest types are subdivided within the group of dwarf shrub-green moss birch forests: dwarf shrub-green moss birch forests and bilberry-green moss birch forests. The first forest type is found in the forests of the northern taiga, and the latter one – in the middle taiga (Fig. 11).
Dwarf shrub-green moss birch forests are usually a derivative post-fire or post-felling version of spruce forests in this section. The ratio of the dominant species of the ground layers and the degree of their development reflect different stages of secondary successions.

Figure 10. Dwarf shrub-green moss birch forest
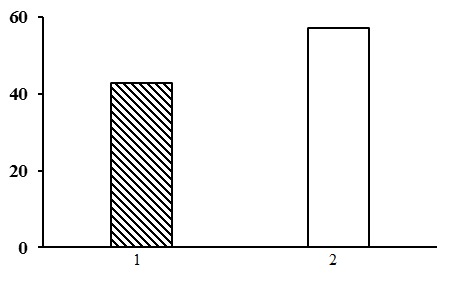
Figure 11. The ratio of forest types in the group of dwarf shrub-green moss birch forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows forest types (1 – dwarf shrub-green moss birch forests, 2 – bilberry-green moss birch forests), the Y-axis shows the percentage
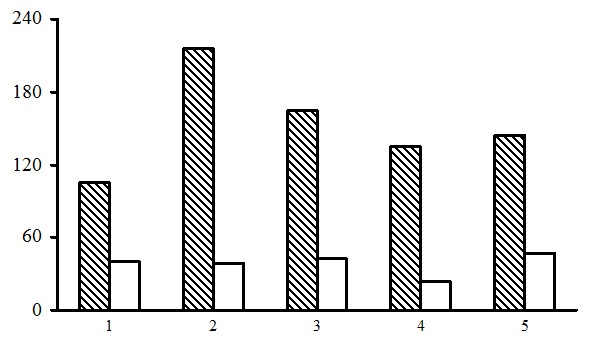
Figure 12. Alpha diversity of the green moss section forests. The shaded columns stand for the total number of species in the forest type group (species richness); unshaded columns – for the average number of species per PLP (species density). The X-axis shows forest types groups (1 – dwarf shrub-green moss birch forests, 2 – dwarf shrub-green moss spruce forests, 3 – small grass-green moss spruce forests, 4 – dwarf shrub-green moss pine forests, 5 – small grass-green moss pine forests), the Y-axis shows the number of plant species (vascular plants, mosses, lichens)
Table 6. Some parameters of the floristic diversity of the dwarf shrub-green moss pine forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
max |
x |
st. dev. |
|
|
Number of species per PLP (vascular plants, mosses and lichens) |
14.0 |
38.0 |
23.6 |
5.0 |
13.0 |
44.0 |
28.7 |
7.7 |
|
Number of vascular plants per PLP |
6.0 |
25.0 |
13.6 |
4.8 |
8.0 |
34.0 |
19.3 |
6.9 |
| Average number of vascular plant species per a PLP plot |
4.8 |
13.5 |
8.2 |
2.7 |
4.5 |
16.8 |
10.3 |
3.7 |
|
Number of moss and lichen species per PLP |
6.0 |
14.0 |
10.0 |
2.2 |
4.0 |
21.0 |
9.4 |
4.7 |
| Average number of moss and lichen species per a PLP plot |
4.0 |
8.8 |
6.4 |
1.3 |
2.8 |
12.0 |
5.3 |
2.4 |
|
Number of PLPs |
25 |
21 |
||||||
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. |
||||||||
Floristic diversity of the forests of the green moss section was estimated using alpha and beta diversity.
Evaluation of alpha diversity. Species density in different forest type groups of the green moss section varies within a rather broad range – from 24 to 47 species (Fig. 12). At the same time the majority of PLPs were found to have 20 to 40 plant species (Tables 6–10). The highest floristic diversity is typical for small grass-green moss pine and spruce forests: up to 80 plant species per PLP. Such forests represent more advanced stages of succession (Morozova et al., 2008). Dwarf shrub-green moss pine forests are the poorest in species density, as most often they result from repetitive fires. The number of plant species per PLP in such forests rarely exceeds 20. Species richness depends primarily on the number of PLPs belonging to a certain group of forest types, but generally it correlates with species density.
Evaluation of beta diversity. Beta diversity (Tables 11 and 12) within the green moss section is assessed using Jaccard index and Whittaker index. At pair-wise comparison of forest type groups of the green moss section Jaccard similarity index varies in the range from 0.2 to 0.6. Dwarf shrub-green moss pine and spruce forests demonstrate the biggest similarity (0.6), so do small grass-green moss pine and spruce forests (0.6); the lowest similarity is found between dwarf shrub-green moss birch forests and small grass-green moss spruce forests (0.2). The value of Whittaker’s index (10.2) here is significantly higher than for different groups of forest types within the section (2.7–5.9).
Table 7. Some parameters of the floristic diversity of the small grass-green moss pine forests
|
Parameters |
Forest area |
|||
|
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
|
|
Number of species per PLP (vascular plants, mosses and lichens) |
38.0 |
81.0 |
53.2 |
15.9 |
| Number of vascular plants per PLP |
32.0 |
73.0 |
46.1 |
14.6 |
| Average number of vascular plant species per a PLP plot |
16.3 |
35.3 |
22.6 |
6.6 |
|
Number of moss and lichen species per PLP |
4.0 |
12.0 |
7.1 |
2.2 |
| Average number of moss and lichen species per a PLP plot |
2.5 |
6.0 |
3.7 |
1.1 |
| Number of PLPs |
9 |
|||
|
Comment. Legend – see Table 6. |
||||
Table 8. Some parameters of the floristic diversity of the dwarf shrub-green moss spruce forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
max |
x |
st. dev. |
|
| Number of species per PLP (vascular plants, mosses and lichens) |
23.0 |
55.0 |
34.4 |
8.9 |
22.0 |
62.0 |
40.1 |
10.4 |
|
Number of vascular plants per PLP |
10.0 |
30.0 |
18.0 |
6.0 |
5.0 |
38.0 |
24.1 |
9.4 |
| Average number of vascular plant species per a PLP plot |
6.3 |
15.0 |
10.3 |
2.8 |
3.5 |
20.8 |
13.1 |
5.2 |
| Number of moss and lichen species per PLP |
9.0 |
25.0 |
16.4 |
5.6 |
6.0 |
36.0 |
15.9 |
7.8 |
| Average number of moss and lichen species per a PLP plot |
6.5 |
12.3 |
8.9 |
2.0 |
2.0 |
22.3 |
9.0 |
4.8 |
|
Number of PLPs |
11 |
16 |
||||||
|
Comment. Legend – see Table 6. |
||||||||
Table 9. Some parameters of the floristic diversity of the small-grass-green moss spruce forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
|
Number of species per PLP (vascular plants, mosses and lichens) |
45.0 |
19.0 |
76.0 |
46.0 |
20.5 |
| Number of vascular plants per PLP |
34.0 |
14.0 |
55.0 |
34.1 |
14.6 |
| Average number of vascular plant species per a PLP plot |
19.5 |
7.8 |
31.8 |
17.5 |
7.8 |
|
Number of moss and lichen species per PLP |
11.0 |
2.0 |
25.0 |
11.9 |
7.9 |
| Average number of moss and lichen species per a PLP plot |
5.5 |
1.0 |
17.3 |
6.4 |
5.4 |
| Number of PLPs |
1 |
9 |
|||
|
Comment. Legend – see Table 6. |
|||||
Ecological-coenotic structure of forests. The results of the floristic list–based calculation of the ecological-coenotic structure of species diversity have shown that the forests of the green moss section have a large number of ecological-coenotic plant groups (Fig. 13). The section is dominated by forest species (boreal, nemoral, nitrophilous and pine-forest) of all forms of life: trees, shrubs, dwarf shrubs and grasses. The share of meadow and forest-edge plants is quite high. This is due to the fact that most of the forests of the green moss section are pine forests with the canopy transmitting enough light for the light-demanding plants of open habitats to exist. The share of nitrophilous species, indicating of an advanced successional status of communities, is low in the studied forests (Zaugolnova, Morozova, 2004).
Table 10. Some parameters of the floristic diversity of the dwarf shrub-green moss birch forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
max |
x |
st. dev. |
|
| Number of species per PLP (vascular plants, mosses and lichens) |
27.0 |
57.0 |
40.8 |
15.4 |
33.0 |
45.0 |
39.3 |
6.1 |
|
Number of vascular plants per PLP |
18.0 |
34.0 |
25.2 |
8.3 |
19.0 |
31.0 |
24.8 |
5.1 |
|
Average number of vascular plant species per a PLP plot |
13.3 |
17.3 |
15.4 |
2.0 |
10.8 |
16.8 |
13.9 |
2.6 |
|
Number of moss and lichen species per PLP |
9.0 |
23.0 |
15.6 |
7.1 |
10.0 |
18.0 |
14.5 |
3.4 |
|
Average number of moss and lichen species per a PLP plot |
5.3 |
11.3 |
8.7 |
3.2 |
6.8 |
9.5 |
8.2 |
1.2 |
|
Number of PLPs |
3 |
4 |
||||||
|
Comment. Legend – see Table 6. |
||||||||
Table 11. Jaccard index values for the forest type groups of the green moss section
|
Forest type group |
БКЗ |
ЕМЗ |
ЕКЗ |
СКЗ |
СМЗ |
|
БКЗ |
1 |
0.2 |
0.4 |
0.4 |
0.3 |
|
ЕМЗ |
— |
1 |
0.3 |
0.5 |
0.6 |
|
ЕКЗ |
— |
— |
1 |
0.6 |
0.5 |
|
СКЗ |
— |
— |
— |
1 |
0.4 |
|
СМЗ |
— |
— |
— |
— |
1 |
| Comment. БКЗ – dwarf shrub—green moss birch forests, ЕКЗ – dwarf shrub—green moss spruce forests, ЕМЗ – small grass—green moss spruce forests, СКЗ – dwarf shrub—green moss pine forests, СМЗ – small grass—green moss pine forests. | |||||
Table 12. Values of beta diversity for the forests of the green moss section
|
Forest type group |
БКЗ |
ЕМЗ |
ЕКЗ |
СКЗ |
СМЗ |
For the section |
|
Average species density |
105 |
215 |
164 |
135 |
144 |
327 |
|
Total number of species |
40 |
39 |
43 |
24 |
47 |
33 |
| Whittaker index (W) |
2.7 |
5.7 |
3.9 |
5.9 |
3.1 |
10.2 |
| Comment. БКЗ – dwarf shrub—green moss birch forests, ЕКЗ – dwarf shrub—green moss spruce forests, ЕМЗ – small grass—green moss spruce forests, СКЗ – dwarf shrub—green moss pine forests, СМЗ – small grass—green moss pine forests. | ||||||
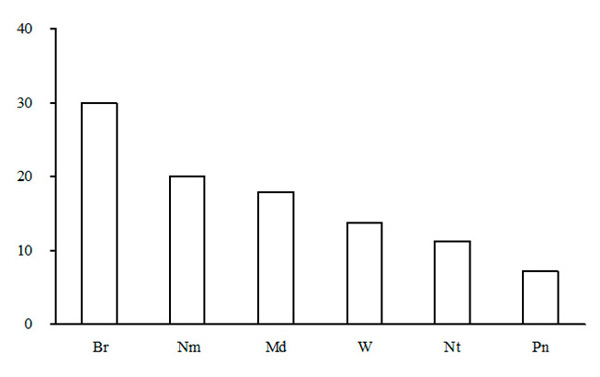
Figure 13. Ecological-coenotic structure of forests of the green moss section. The X-axis shows ecological-coenotic plant groups (Br – boreal, Nm – nemoral, Pn – pine-forest, Md – meadow and meadow-forest edge, Nt – nitrophilous, W – wetland),
the Y-axis shows the percentage
The forests of the green moss section are very common both in the northern and the middle taiga. They are mostly various monodominant pine forests with a relatively high tree canopy density, the predominance of one or two types of dominants in the grass-dwarf shrub layer, and well-developed moss cover of boreal green mosses. In the long-term absence of fires or felling, spruce understorey can enter the tree layer, and pine forests with spruce are replaced by spruce forests with singular pine trees. At the same time, spruce forests quite often demonstrate post-fire origin as well (Volkov, 2008; Zaugolnova, Morozova, 2004). Birch forests usually represent a post-fire or post-felling derivative version of pine and spruce forests of this section, which is proven by the floristic similarity of these forests.
Lichenous section (cladinosa)
Classification and characteristics of forests. Communities of the lichenous section are not common in the studied forests: of all the established PLPs only 8% belong to the lichenous section. The majority of them was found in the forests of the northern taiga. They are different pine forests with a well-developed moss-lichen cover: as a rule, the coverage of the layer is at least 50%. The lichen coverage exceeds or is almost equal to that of mosses.
Only two forest types groups are represented in the lichenous section: lichen pine forests and lichen-green moss pine forests. Their share is equal. Let us look at the groups of forest types of the lichenous section.
Lichen pine forests (geobotanical description – see Table 13) were found only in the forests of the northern taiga. The tree layer is dominated by Pinus sylvestris with a small admixture of Betula pubescens (Fig. 14). The understorey and shrubs is usually sparse. A young generation of trees (Betula pubescens, Picea abies) and shrubs (Juniperus communis, Salix caprea, etc.) is observed here. The grass-dwarf shrub layer is codominated by Calluna vulgaris, Vaccinium vitis-idaea, V. myrtillus, Empetrum nigrum, etc. The moss-lichen layer is characterized by the predominance of lichens: Cladonia arbuscula, C. rangiferina, C. stellaris, etc. Mosses are represented by Pleurozium schreberi, Polytrichum commune, Dicranum polysetum, Dicranum scoparium, etc.

Figure 14. Lichen pine forest
Table 13. Some geobotanical parameters of the lichen pine forests
|
Parameters |
Forest area |
|||
|
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
|
|
Stand age, yrs |
30.0 |
60.0 |
40.2 |
12.5 |
|
Stand height, m |
6.0 |
13.0 |
8.5 |
2.8 |
|
Tree canopy density (A), % |
3.8 |
57.5 |
35.5 |
19.2 |
| Density of the layer of shrubs and tree understorey (B), % |
3.5 |
18.0 |
8.2 |
5.9 |
| Total projective cover of the grass layer (C), % |
17.0 |
56.1 |
30.5 |
13.6 |
|
Total projective cover of the moss-lichen layer (D), % |
6.2 |
83.8 |
60.6 |
30.1 |
| Total projective cover of green mosses in the moss-lichen layer, % |
0.2 |
30.9 |
16.0 |
13.1 |
| Total projective cover of lichens in the moss-lichen layer, % |
6.0 |
75.7 |
44.6 |
23.3 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
0.0 |
0.0 |
0.0 |
| Number of PLPs |
6 |
|||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | ||||
Three types of forests are subdivided within the group of lichen pine forests: heather-lichen pine forests, heathberry-cowberry-lichen pine forests and cowberry-lichen pine forests, with the domination of the heather-lichen pine forests (Fig. 15).
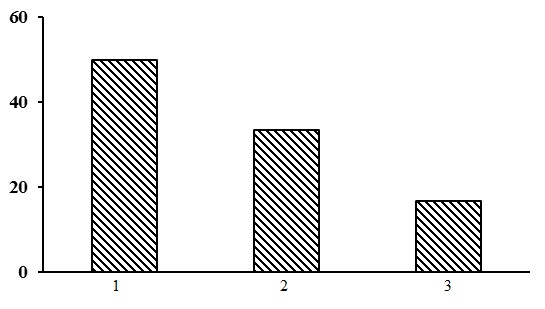
Figure 15. The ratio of forest types in the group of lichen pine forests. The shaded columns stand for the northern taiga. The X-axis shows forest types (1– heather-lichen pine forests, 2 – heathberry-cowberry-lichen pine forests, 3 – cowberry-lichen pine forests), the Y-axis shows the percentage
Green moss-lichen pine forests (geobotanical description – see Table 14). The tree layer consists of Pinus sylvestris with a small admixture of Betula pubescens and Picea abies. The shrubs layer and the understorey are relatively poorly developed (Fig. 16). It consists of a young generation of trees (Pinus sylvestris, Betula pubescens and Picea abies) and shrubs (Sorbus aucuparia, Salix caprea, Juniperus communis, etc.). The grass-dwarf shrub layer is formed by the predominant dwarf shrubs: Vaccinium myrtillus, V. vitis-idaea, Calluna vulgaris, Ledum palustre, Empetrum nigrum, E. hermaphroditum, etc. There are singular boreal grasses: Equisetum sylvaticum, Carex globularis, Avenella flexuosa, etc. The moss-lichen layer consists of lichens (Cladonia arbuscula, C rangiferina, etc.) and green mosses (Pleurozium schreberi, Hylocomiun splendens, etc.).
Three types of forests are subdivided within the group of green moss-lichen pine forests: green moss-lichen heather pine forests, green moss-lichen bilberry pine forests, and green moss-lichen cowberry pine forests. There are only a few PLPs for each forest type. Predominant forests are green moss-lichen heather pine forests, which are found both in the northern (2 PLPs) and middle (1 PLP) taiga (Fig. 17).
Table 14. Some geobotanical parameters of the green moss-lichen pine forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
|
Stand age, yrs |
40.0 |
210.0 |
104.0 |
70.9 |
34.0 |
|
Stand height, m |
6.0 |
19.0 |
13.6 |
5.0 |
11.0 |
|
Tree canopy density (A), % |
18.8 |
42.5 |
33.0 |
8.9 |
40.0 |
| Density of the layer of shrubs and tree understorey (B), % |
2.3 |
15.0 |
6.9 |
4.9 |
2.8 |
| Total projective cover of the grass layer (C), % |
34.7 |
67.1 |
50.2 |
15.1 |
25.5 |
| Total projective cover of the moss-lichen layer (D), % |
72.8 |
95.4 |
87.2 |
8.8 |
91.8 |
| Total projective cover of green mosses in the moss-lichen layer, % |
25.6 |
47.7 |
39.0 |
9.7 |
47.3 |
| Total projective cover of lichens in the moss-lichen layer, % |
27.6 |
51.8 |
39.3 |
10.4 |
44.5 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
32.5 |
8.9 |
14.0 |
0.0 |
|
Number of PLPs |
5 |
1 |
|||
| Comment. Legend – see Table 13. | |||||

Figure 16. Lichen-green moss pine forest
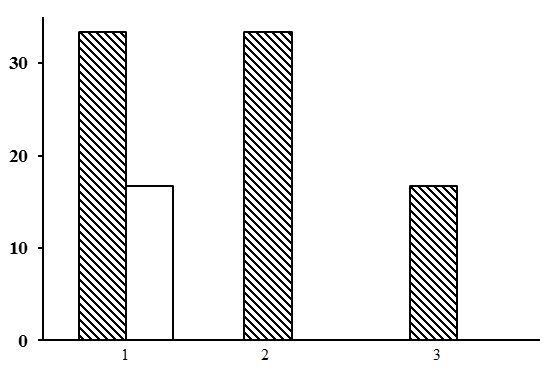
Figure 17. The ratio of forest types in the group of green moss-lichen pine forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows forest types (1 – green moss-lichen heather pine forests, 2 – green moss-lichen cowberry pine forests, 3 – green moss-lichen bilberry pine forests), the Y-axis shows the percentage
Floristic diversity of forests of the lichenous section was evaluated using alpha and beta diversity.
Evaluation of alpha diversity. Species density in different forest type groups of the lichenous section varies within a small range: from 22 to 29 species (Fig. 18). At the same time at the majority of PLPs from 20 to 30 plant species were found (Tables 15 and 16). Lichen pine forests had the lowest values of floristic diversity. For example, in the heather-lichen pine forests, which predominate in this group of forest types, there are only 16 to 19 plant species per PLP. The floristic diversity of the green moss-lichen pine forests is higher than that of the lichen pine forests. For example, in the green moss-lichen cowberry and bilberry pine forests there are up to 36 species per PLP.
Evaluation of beta diversity. Beta diversity (Tables 17 and 18) within the lichenous section is estimated using the value of Jaccard index and Whittaker’s index. When comparing lichen pine forests and green moss-lichen pine forests, the value of Jaccard index of similarity was high (0.7). This means there is a significant floristic similarity of these groups of forest types. Whittaker’s index value (3.3) for the lichenous section is very close to the equivalent parameter for the groups of forest types within a section: 2.4 (lichen pine forests) and 2.5 (green moss-lichen pine forests).
Table 15. Some parameters of the floristic diversity of lichen pine forests
|
Parameters |
Forest area |
|||
|
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
|
|
Number of species per PLP (vascular plants, mosses and lichens) |
16.0 |
29.0 |
21.5 |
4.5 |
|
Number of vascular plants per PLP |
6.0 |
16.0 |
11.0 |
3.2 |
| Average number of vascular plant species per a PLP plot |
4.3 |
9.5 |
7.6 |
2.0 |
|
Number of moss and lichen species per PLP |
5.0 |
18.0 |
10.5 |
4.7 |
| Average number of moss and lichen species per a PLP plot |
3.3 |
14.3 |
7.5 |
4.0 |
|
Number of PLPs |
6 |
|||
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. |
||||
Table 16. Some geobotanical parameters of the green moss-lichen pine forests
|
Parameters |
Forest area |
Northern taiga |
|||
|
Northern taiga |
|||||
|
min |
max |
x |
st. dev. |
||
|
Number of species per PLP (vascular plants, mosses and lichens) |
16.0 |
37.0 |
28.2 |
9.9 |
30.0 |
|
Number of vascular plants per PLP |
7.0 |
20.0 |
12.6 |
5.0 |
13.0 |
| Average number of vascular plant species per a PLP plot |
5.3 |
12.3 |
8.6 |
3.1 |
7.0 |
|
Number of moss and lichen species per PLP |
9.0 |
24.0 |
15.6 |
6.4 |
17.0 |
| Average number of moss and lichen species per a PLP plot |
6.8 |
12.3 |
9.3 |
2.6 |
9.0 |
|
Number of PLPs |
5 |
1 |
|||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | |||||
Table 17. Values of Jaccard index for the forest type groups of the lichenous section
|
Forest type group |
СЛ |
СЗЛ |
|
СЛ |
1 |
0.7 |
|
СЗЛ |
— |
1 |
| Comment. СЛ – lichen pine forests, СЗЛ – green moss-lichen pine forests | ||
Table 18. Values of beta diversity of forests of the lichenous section
|
Forest type group |
СЛ |
СЗЛ |
For the section |
|
| Total number of the species |
51.0 |
70.0 |
80 |
|
|
Average species density |
22.0 |
29.0 |
25 |
|
| Whittaker’s index (W) |
2.4 |
2.5 |
3.3 |
|
| Comment. СЛ – lichen pine forests, СЗЛ – green moss-lichen pine forests | ||||
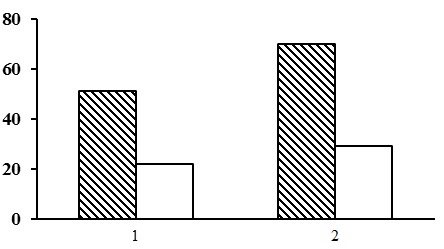
Figure 18. Alpha diversity of forests of the lichenous section. The shaded columns stand for the total number of species in the forest type group (species richness); the unshaded columns – for the average number of species per PLP (species density). The X-axis shows groups of forest types (1– lichen pine forests, 2 – green moss-lichen pine forests), the Y-axis – the number of plant species (vascular plants, mosses, lichens)
Ecological-coenotic structure of forests. The results of the floristic list–based calculation of the ecological-coenotic structure of species diversity have shown that the forests of the lichenous section have a fairly diverse set of ecological-coenotic plant groups (Fig. 19). It contains boreal forest species of all life forms: trees, shrubs, dwarf shrubs and grasses. High share of wetland plants is due to the fact that pine forests of the lichenous section, usually grow on ridges among the marshes. The share of meadow and forest-edge plants is insignificant, which is typical for taiga forests. It is important to note that there are no nitrophilous plant species in the studied forests.
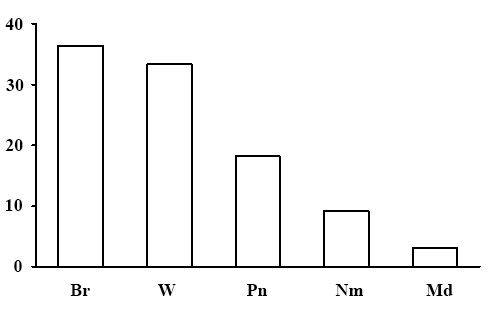
Figure 19. Ecological-coenotic forest structure of the lichenous section. The X-axis shows ecological-coenotic plant groups (Br – boreal, Nm – nemoral, Pn – pine-forest, Nt – nitrophilous, W – wetland, Md – meadow and meadow-forest edge), the Y-axis shows the percentage
Lichen forests are not common in the study area. They are mainly found in the northern taiga. These are various monodominant pine forests (lichen and green moss-lichen) with low values of species density and species richness. Lichen pine forests usually are of pyrogenic origin. Green moss-lichen pine forests are a transitional stage of post-fire secondary successions: from lichen pine forests to green moss pine forests. Repetitive fires, however, often prevent secondary successions from going on. (Volkov, 2008). The fact that forests of the lichenous section are at the initial stage of succession is confirmed by low floristic diversity of communities, poorly developed layer of shrubs and understorey of trees (underbrush), the predominance of one or two dominant species in the grass-dwarf shrub layer and the absence of rare species.
Sphagnous section (sphagnosa)
Classification and characteristics of forests. The communities of the sphagnous section are relatively rare in the forests under study: of all the established PLPs only 16% belong to the sphagnous section. Of these, 20% are the forests of the northern taiga, and 80% – the forests of the middle taiga, being mainly various pine and spruce forests. The forests of the sphagnous section have a well-developed moss cover: the coverage of mosses is usually at least 50%. The coverage is dominated by sphagnum mosses, sometimes combined with Polytrichum commune. The grass-dwarf shrub layer is usually dominated by boreal species (grasses and dwarf shrubs), whereas wetland species as a rule show low abundance.
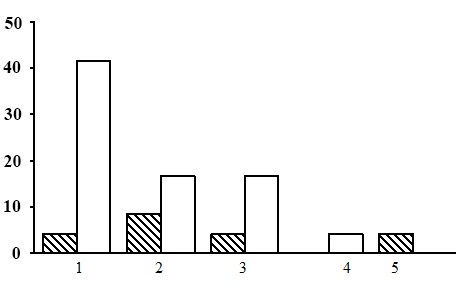
Figure 20. Ratio of PLPs by groups of forest types. Sphagnous section. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows groups of forest types (1 – long-stem moss-sphagnum spruce forests, 2 – dwarf shrub-sphagnum pine forests, 3 – long-stem moss-sphagnum pine forests, 4 – long-stem moss-sphagnum birch forests,5 – eumesotrophic grass-sphagnum spruce forests), the Y-axis shows percentage
The sphagnous section includes the forests belonging to five groups of forest types: long-stem moss-sphagnum birch forests, dwarf shrub-sphagnum pine forests, long-stem moss-sphagnum pine forests, long-stem moss-sphagnum spruce forests, and eumesotrophic grass-sphagnum spruce forests. Among the listed groups of forest types the most common are long-stem moss-sphagnum spruce forests (Fig. 20). They account for about 50% of the PLPs belonging to the sphagnous section. The second most common are dwarf shrub-sphagnum pine forests and long-stem moss-sphagnum pine forests. The share of long-stem moss-sphagnum birch forests and eumesotrophic grass-sphagnum spruce forests is insignificant. Let us consider the groups of forest types of the sphagnous section.
Long-stem moss-sphagnum spruce forests (geobotanical description – see Table 19). Picea abies dominates in the tree layer, Betula pubescens, Pinus sylvestris, etc. can be found as an admixture (Fig. 21). The shrubs layer and the understorey consist of dominant Picea abies; there are singular plants of Salix caprea, Alnus incana, Betula pubescens, Pinus sylvestris, Sorbus aucuparia etc. The grass-dwarf shrub layer is dominated by boreal dwarf shrubs: Vaccinium myrtillus and Vaccinium vitis-idaea; while Rubus chamaemorus, Equisetum sylvaticum, Carex globularis and others occur with high constancy but are of low abundance. Viola epipsila, Equisetum palustre and other species indicative of waterlogging were noticed. Sphagnum mosses dominate in the moss-lichen layer: Sphagnum girgensohnii, S. angustifolium, etc. Polytrichum commune, Hylocomium splendens, and Pleurozium schreberi are common among the green mosses. The literature describes long stem moss-sphagnum spruce forests with the predominance of Polytrichum commune (Korchagin, 1940). It is associated with fires: after the trees death, evaporation drops dramatically, and the development of the haircap moss-based coverage further deteriorates of the hydrological regime.

Figure 21. Long stem moss-sphagnum spruce forest
Two types of forest are subdivided within the group of long stem moss-sphagnum spruce forests: bilberry-sphagnum spruce forests and bilberry-long stem moss-sphagnum spruce forests (Fig. 22). Predominant are the bilberry-long-stem moss-sphagnum spruce forests, which are found only in the middle taiga.
Long stem moss-sphagnum spruce forests grow on the flat depressions between the ridges in floodplain landscapes as well as in the heads of small rivers and mesorelief depressions.
Long-stem-sphagnum pine forests (geobotanical description – see Table 20) are forests with the tree layer dominated by Pinus sylvestris. Betula pubescens quite often shows high abundance. The shrubs layer and the understorey consist of adult shrubs (Juniperus communis, Sorbus aucuparia) and young generation of trees (Betula pubescens, Pinus sylvestris). The grass-dwarf shrub layer in both the northern and middle taiga is dominated by Vaccinium myrtillus and Rubus chamaemorus. The high constancy species are Vaccinium uliginosum, Ledum palustre, Vaccinium vitis-idaea, Avenella flexuosa, Carex globularis. The share of boreal small grasses increases from the northern to the middle taiga; in addition, Empetrum hermaphroditum is found in the northern taiga. The moss-lichen layer is dominated by sphagnum (Sphagnum angustifolium) and green (Pleurozium schreberi) mosses. At the same time other types of sphagnum mosses: Sphagnum russowii, S. magellanicum, etc., and admixture of lichens is found there.
Table 19. Some geobotanical parameters of the long-stem moss-sphagnum spruce forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
|
Stand age, yrs |
130.0 |
75.0 |
120.0 |
96.0 |
14.1 |
|
Stand height, m |
19.0 |
13.0 |
25.0 |
21.2 |
4.0 |
|
Tree canopy density (A), % |
35.0 |
21.8 |
74.3 |
49.5 |
17.9 |
| Density of the layer of shrubs and tree understorey (B), % |
10.0 |
4.6 |
22.6 |
11.6 |
5.7 |
| Total projective cover of the grass layer (C), % |
72.4 |
17.1 |
44.2 |
27.1 |
8.5 |
| Total projective cover of the moss-lichen layer (D), % |
83.1 |
23.1 |
92.8 |
67.2 |
22.1 |
| Total projective cover of green mosses in the moss-lichen layer, % |
18.1 |
0.1 |
40.3 |
16.2 |
15.2 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
0.0 |
0.8 |
0.1 |
0.2 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
65.0 |
23.0 |
77.0 |
51.0 |
18.0 |
|
Number of PLPs |
1 |
10 |
|||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | |||||
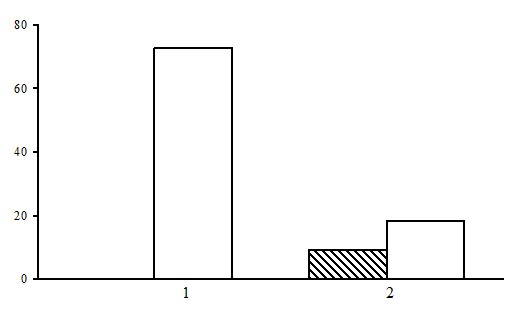
Figure 22. The ratio of forest types in the group of long-stem moss-sphagnum spruce forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows forest types (1 – bilberry-long-stem moss-sphagnum spruce forests, 2 – bilberry-sphagnum spruce forests), the Y-axis shows the percentage
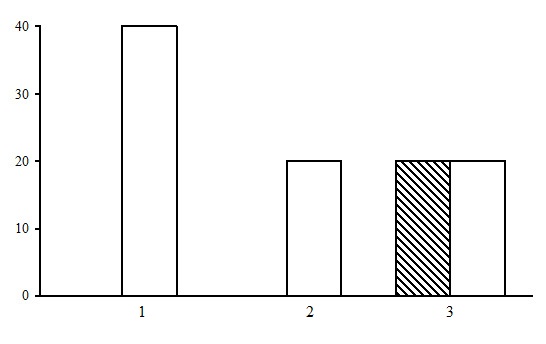
Figure 23. The ratio of forest types in the group of long-stem moss-sphagnum pine forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-line shows forest types (1 – bilberry-sphagnum pine forests, 2 – cloudberry-sphagnum pine forests, СЧЗС – bilberry-green moss-sphagnum pine forests), the Y-axis shows the percentage
Three types of forests are subdivided within the group of long-stem moss-sphagnum pine forests: cloudberry-sphagnum pine forests, bilberry-green moss-sphagnum pine forests, and bilberry-sphagnum pine forests. The predominant type is bilberry-sphagnum pine forests, which are found only in the middle taiga (Fig. 23). Bilberry-green moss-sphagnum pine forests are found both in the northern and in the middle taiga. They represent the initial stages of waterlogging of dwarf shrub-green moss forests.
Long-stem moss-sphagnum pine forests grow on poorly drained areas of the plains, on the smooth slopes of ridges with peaty and peaty-gley soils.
Dwarf shrub-sphagnum pine forests (geobotanical description – see Table 21). The tree layer is formed by predominant Pinus sylvestris, with Betula pubescens, Picea abies etc. can occur as an admixture (Fig. 24). The shrubs layer and the understorey are dominated by the younger generation of trees that form the forest stand. The grass-dwarf shrub layer is characterized by the constant presence of Vaccinium uliginosum, Vaccinium myrtillus, Ledum palustre, Carex globularis, Eriophorum vaginatum, Rubus chamaemorus, Oxycoccus palustris, etc. Oligotrophic species most often dominate among them: Vaccinium uliginosum and Ledum palustre. The predominance of sphagnum mosses is typical for the moss-lichen layer: Sphagnum angustifolium, S. fuscum, S. magellanicum. The forests of this group are characterized by a hummocky microrelief and a mosaic structure of the ground layers.
Table 20. Some geobotanical parameters of the long-stem moss-sphagnum pine forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
|
Stand age, yrs |
130.0 |
65.0 |
130.0 |
91.0 |
24.1 |
| Stand height, m |
14.0 |
7.0 |
26.0 |
17.8 |
7.4 |
|
Tree canopy density (A), % |
18.5 |
17.5 |
100.0 |
49.2 |
34.5 |
| Density of the layer of shrubs and tree understorey (B), % |
4.5 |
0.8 |
16.3 |
8.4 |
6.3 |
| Total projective cover of the grass layer (C), % |
46.6 |
28.4 |
55.4 |
44.2 |
11.3 |
| Total projective cover of the moss-lichen layer (D), % |
79.3 |
26.9 |
99.5 |
57.2 |
31.2 |
| Total projective cover of green mosses in the moss-lichen layer, % |
21.0 |
1.0 |
26.4 |
13.4 |
11.1 |
| Total projective cover of lichens in the moss-lichen layer, % |
1.0 |
0.0 |
1.0 |
0.2 |
0.4 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
57.3 |
17.0 |
98.5 |
43.6 |
34.7 |
|
Number of PLPs |
1 |
4 |
|||
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. |
|||||
Two types of forest are subdivided within the group of dwarf shrub-sphagnum pine forests: dwarf shrub-sphagnum pine forests and ledum-sphagnum pine forests. The predominant type is dwarf shrub-sphagnum pine forests, which are found both in the northern and the middle taiga (Fig. 25). Ledum-sphagnum pine forests were found only in the middle taiga.
Dwarf shrub-sphagnum pine forests are usually located in poorly drained relief depressions or on the margins of oligotrophic bogs.
Table 21. Some geobotanical parameters of the dwarf shrub-sphagnum pine forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
max |
x |
st. dev. |
|
|
Stand age, yrs |
80.0 | 300.0 | 190.0 | 155.6 | 30.0 | 100.0 | 65.0 | 31.1 |
|
Stand height, m |
10.0 | 11.0 | 10.5 | 0.7 | 6.0 | 14.0 | 8.5 | 3.7 |
|
Tree canopy density (A), % |
15.0 | 32.5 | 23.8 | 12.4 | 7.5 | 55.0 | 23.1 | 22.1 |
| Density of the layer of shrubs and tree understorey (B), % | 13.5 | 16.3 | 14.9 | 1.9 | 1.8 | 31.3 | 12.9 | 12.8 |
| Total projective cover of the grass layer (C), % | 41.7 | 67.8 | 54.7 | 18.4 | 17.3 | 61.7 | 41.7 | 18.9 |
| Total projective cover of the moss-lichen layer (D), % | 59.4 | 90.0 | 74.7 | 21.6 | 94.0 | 98.9 | 96.9 | 2.3 |
| Total projective cover of green mosses in the moss-lichen layer, % | 2.5 | 6.1 | 4.3 | 2.6 | 0.1 | 11.7 | 4.4 | 5.4 |
| Total projective cover of lichens in the moss-lichen layer, % | 0.1 | 0.8 | 0.4 | 0.5 | 0.0 | 2.7 | 0.7 | 1.3 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % | 52.5 | 87.5 | 70.0 | 24.7 | 84.4 | 98.8 | 91.8 | 7.6 |
|
Number of PLPs |
2 |
4 |
||||||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | ||||||||

Figure 24. Dwarf shrub-sphagnum pine forest
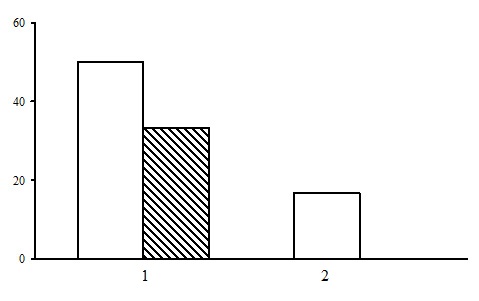
Figure 25. The ratio of forest types in the group of dwarf shrub-sphagnum pine forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows types of forests (1 – dwarf shrub-sphagnum pine forests, 2 – ledum-sphagnum pine forests), the Y-axis shows the percentage
Eumesotrophic grass-sphagnum spruce forests (geobotanical description – see Table 22) are represented by only one PLP (grass-cowberry-sphagnum spruce forest), which is located in the northern taiga. The trees layer is formed by predominant Picea abies with an admixture of Betula pubescens. The shrubs layer and the understorey have a low abundance of Betula pubescens, Juniperus communis, Sorbus aucuparia, Rosa acicularis, etc. The grass-shrub layer is characterized by the presence of boreal dwarf shrubs (Vaccinium vitis-idaea), eutrophic and mesotrophic grasses. In the moss-lichen cover species of the genus Sphagnum are constant. Green boreal mosses (Hylocomium splendens, Pleurozium schreberi) also occur, but with low abundance.
Table 22. Some geobotanical parameters of the eumesotrophic grass-sphagnum spruce forest and the long-stem moss-sphagnum birch forest
|
Parameters |
Forest type |
|
|
ЕТБрС |
БХС |
|
|
Stand age, yrs |
80.0 |
70.0 |
|
Stand height, m |
15.0 |
20.0 |
|
Tree canopy density (A), % |
46.3 |
81.3 |
| Density of the layer of shrubs and tree understorey (B), % |
9.0 |
11.5 |
| Total projective cover of the grass layer (C), % |
35.5 |
44.2 |
| Total projective cover of the moss-lichen layer (D), % |
79.7 |
69.8 |
| Total projective cover of green mosses in the moss-lichen layer, % |
12.7 |
15.0 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
0.0 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
67.0 |
54.9 |
|
Number of PLPs |
1 |
1 |
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. ЕТБрС – grass—cowberry sphagnum spruce forest, БХС – horsetail—sphagnum birch forest. |
||
Long-stem moss-sphagnum birch forests (geobotanical description – see Table 22) are represented by only one PLP (horsetail-sphagnum birch forest), which is located in the middle taiga. The tree layer is formed by Betula pendula, B. pubescens, and Populus tremula. The shrubs layer and the understorey consist of a young generation of trees (Picea abies) and adult shrubs (Sorbus aucuparia). The grass-dwarf shrub layer is dominated by Equisetum sylvaticum. Vaccinium myrtillus, Gymnocarpium dryopteris, Oxalis acetosella, etc. are found as admixtures. Sphagnum mosses dominate in the moss-lichen layer: Sphagnum girgensohnii.
Floristic diversity of forests of the sphagnous section was estimated using alpha and beta diversity indicators.
Evaluation of alpha diversity. Species density in different forest types groups of the sphagnous section is virtually the same: from 28 to 29 species (Fig. 26). On the majority of PLPs about 30 plant species were identified (Tables 23–26). Long stem moss- and dwarf shrub-sphagnum pine forests show the lowest values of floristic diversity, whereas long stem moss-sphagnum spruce forests are the most diverse.
Evaluation of beta diversity. Beta diversity (Tables 27 and 28) within the sphagnous section is estimated using Jaccard index and Whittaker’s index.
In the pair-wise comparison of forest type groups of the sphagnous section, the value of Jaccard index of similarity varies from 0.3 to 0.6. Most similar in floristic composition were long-stem moss-sphagnum pine and spruce forests (0.6), as well as long-stem moss-sphagnum spruce and birch forests (0.5); least similar were long-stem moss-sphagnum birch forests and eumesotrophic grass-sphagnum spruce forests (0.2). Whittaker’s index value (5.7) for the sphagnous section is higher than for the forest types groups within the section (2.6–4.0).
Table 23. Some parameters of floristic diversity of the long-stem moss spruce forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
|
Number of species per PLP (vascular plants, mosses and lichens) |
48.0 |
20.0 |
39.0 |
29.9 |
7.5 |
|
Number of vascular plants per PLP |
41.0 |
8.0 |
31.0 |
21.5 |
7.5 |
| Average number of vascular plant species per a PLP plot |
23.0 |
4.3 |
15.5 |
10.4 |
3.9 |
|
Number of mosses and lichens species per PLP |
7.0 |
5.0 |
17.0 |
8.4 |
3.6 |
| Average number of moss and lichen species per a PLP plot |
4.5 |
3.3 |
9.5 |
5.0 |
2.0 |
|
Number of PLPs |
1 |
10 |
|||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | |||||
Table 24. Some parameters of the floristic diversity of the long-stem moss-sphagnum pine forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
| Number of species per PLP (vascular plants, mosses and lichens) |
28.0 |
25.0 |
41.0 |
30.2 |
6.5 |
|
Number of vascular plants per PLP |
14.0 |
14.0 |
34.0 |
20.6 |
7.7 |
| Average number of vascular plant species per a PLP plot |
8.8 |
8.8 |
19.3 |
12.2 |
4.3 |
|
Number of mosses and lichens species per PLP |
14.0 |
7.0 |
14.0 |
9.6 |
3.2 |
| Average number of moss and lichen species per a PLP plot |
7.3 |
3.5 |
7.3 |
5.5 |
1.6 |
| Number of PLPs |
1 |
4 |
|||
|
Comment. Legend – see Table 23. |
|||||
Table 25. Some parameters of the floristic diversity of the dwarf shrub-sphagnum pine forests
|
Parameters |
Forest area |
|||||||
|
Northern taiga |
Middle taiga |
|||||||
|
min |
max |
x |
st. dev. |
min |
max |
x |
st. dev. |
|
| Number of species per PLP (vascular plants, mosses and lichens) |
33.0 |
38.0 |
35.5 |
3.5 |
13.0 |
38.0 |
24.5 |
33.0 |
|
Number of vascular plants per PLP |
21.0 |
24.0 |
22.5 |
2.1 |
8.0 |
15.0 |
11.8 |
21.0 |
| Average number of vascular plant species per a PLP plot |
14.3 |
16.8 |
15.5 |
1.8 |
6.3 |
11.5 |
8.6 |
14.3 |
| Number of moss and lichen species per PLP |
12.0 |
14.0 |
13.0 |
1.4 |
4.0 |
23.0 |
12.8 |
12.0 |
| Average number of moss and lichen species per a PLP plot |
5.0 |
6.0 |
5.5 |
0.7 |
2.8 |
10.8 |
7.0 |
5.0 |
|
Number of PLPs |
2 |
4 |
||||||
|
Comment. Legend – see Table 23. |
||||||||
Table 26. Some parameters of the floristic diversity of the eumesotrophic grass-sphagnum spruce forest and the long-stem moss-sphagnum birch forest
|
Parameters |
Forest type |
|
|
ЕТБрС |
БХС |
|
|
Number of species per PLP (vascular plants, mosses and lichens) |
51.0 |
61.0 |
|
Number of vascular plants per PLP |
39.0 |
44.0 |
| Average number of vascular plant species per a PLP plot |
18.0 |
20.0 |
|
Number of mosses and lichens species per PLP |
12.0 |
17.0 |
| Average number of moss and lichen species per a PLP plot |
6.0 |
12.0 |
|
Number of PLPs |
1 |
1 |
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. ЕТБрС – grass—cowberry sphagnum spruce forest, БХС – horsetail—sphagnum birch forest. |
||
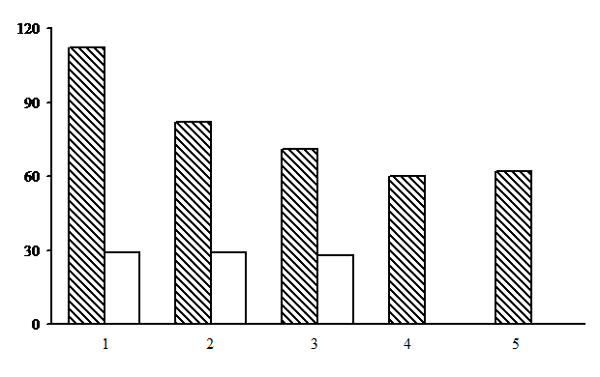
Figure 26. Alpha diversity of forests of the sphagnous section. The shaded columns stand for the total number of species in the forest type group (species richness); the unshaded columns – for the average number of species per PLP (species density). The X-axis shows groups of forest types (1 – long-stem moss-sphagnum spruce forests, 2 – long-stem moss-sphagnum pine forests, 3 – dwarf shrub-sphagnum pine forests, 4 – eumesotrophic grass-sphagnum spruce forests, 5 – long-stem moss-sphagnum birch forests), the Y-axis – the number of plant species (vascular plants, mosses, lichens)
Ecological-coenotic structure of forests. The results of the floristic list–based calculation of the ecological-coenotic structure of species diversity have shown that the forests of the sphagnous section have a fairly diverse set of ecological-coenotic plant groups (Fig. 27). Boreal forest species of all life forms are predominant: trees, shrubs, dwarf shrubs and grasses. The share of marsh plants is high, which is typical for wetland forests. Nitrophilous species were found. The share of meadow and forest-edge plants is insignificant.
The sphagnum-section forests are relatively rare in the study area. They are mainly found in the middle taiga. Long-stem moss-sphagnum pine and spruce forests are predominant there. They are considered to represent the initial stages of waterlogging of the oligotrophic series. The sequence of the change of dominants and codominants with increased waterlogging: Vaccinium myrtillus – Equisetum sylvaticum + Rubus chamaemorus – Carex globularis (Morozova et al., 2008). There are primary (multiple-aged) and derivative spruce forests. The latter arose due to the preservation of the understorey and thinners after fellings of the middle of the 20th century (Martynenko, 1999). Long-stem moss birch forests grow in clear cur areas and burnt areas instead of green moss spruce and pine forests or instead of long-stem moss spruce forests. It is proven by the similarity of their species composition.
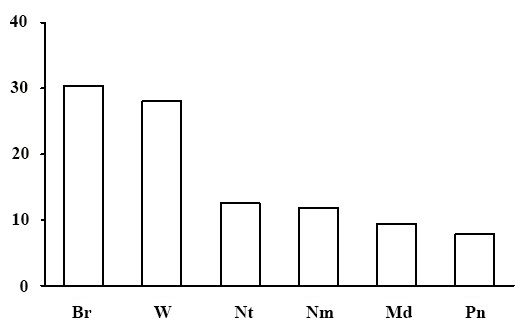
Figure 27. Ecological-coenotic structure of the sphagnous section forests. The X-axis shows ecological-coenotic plant groups (Br – boreal, Nm – nemoral, Pn – pine-forest, Md – meadow and meadow-forest edge, Nt – nitrophilous, W – wetland), the Y-axis shows the percentage
Table 27. Jaccard index for groups of forest types of the sphagnous section
|
Forest type group |
БДС |
СКС |
СДС |
ЕДС |
ЕТС |
|
БДС |
1 |
0.3 |
0.4 |
0.5 |
0.2 |
|
СКС |
— |
1 |
0.3 |
0.4 |
0.3 |
|
СДС |
— |
— |
1 |
0.6 |
0.3 |
|
ЕДС |
— |
— |
— |
1 |
0.4 |
|
ЕТС |
— |
— |
— |
— |
1 |
| Comment. БДС – long—stem moss—sphagnum birch forests, СКС – dwarf shrub—sphagnum pine forests, СДС – long—stem moss—sphagnum pine forests, ЕДС – long—stem moss—sphagnum spruce forests, ЕТС – eumesotrophic grass—sphagnum spruce forests | |||||
Table 28. Beta diversity values of forests of the sphagnous section
|
Forest type group |
ЕДС |
СДС |
СКС |
ЕТС |
БДС |
For the section |
||
|
Total number of species |
112.0 |
82.0 |
71.0 |
60.0 |
62.0 |
159 |
||
|
Average species density |
29.0 |
29.0 |
28.0 |
— |
— |
29 |
||
| Whittaker’s index (W) |
4.0 |
2.9 |
2.6 |
— |
— |
5.7 |
||
| Comment. БДС – long—stem moss—sphagnum birch forests, СКС – dwarf shrub—sphagnum pine forests, СДС – long—stem moss—sphagnum pine forests, ЕДС – long—stem moss—sphagnum spruce forests, ЕТС – eumesotrophic grass—sphagnum spruce forests | ||||||||
Grass (herbosa) and grass-marsh section (uliginosoherbosa)
Classification and characteristics of forests. The grass and grass-marsh sections are analyzed together for several reasons. Firstly, these forests are similar in floristic composition and environmental conditions; and secondly, the grass-marsh section accounts for only one PLP. The communities belonging to the grass and grass-marsh sections are not often found in the studied forests: of all the established PLPs the grass and grass-marsh sections account for only about 10%. There are 15% of the northern taiga forests, and 85% – of the middle taiga forests among them, mainly various birch forests. The forests of the grass section are located on poorly or moderately moist habitats, and the grass-marsh – on excessively moist habitats (water can stay on the surface for a long time). At the same time, both sections are characterized by poorly developed moss and lichen covers (coverage is less than 20%, although in some places it can reach 50%).
The grass and grass-marsh sections include the forests belonging to four forest type groups: small grass birch/aspen forests, grassmarsh birch forests, tall herb birch forests and tall herb spruce forests (Fig. 28). The predominant group is small grass birch/aspen forests (about 60%). The share of other groups of forest types is insignificant. Let us look at the groups of forest types of the grass and grass-marsh sections.
Small grass birch/aspen forests (geobotanical description – see Table 29) show the predominance of Betula pubescens in the tree layer with an admixture of B. pendula, Picea abies, etc. (Fig. 29). The shrubs layer and the understorey consist of a young generation of trees (Picea abies, Betula pubescens) and adult shrubs (Sorbus aucuparia, Juniperus communis, etc.). The grass-dwarf shrub layer is codominated by Vaccinium myrtillus and Equisetum sylvaticum. Gymnocarpium dryopteris, Oxalis acetosella, Trientalis europaea and Maianthemum bifolium are found here with a low abundance but high constancy.
Tall herb (Geranium sylvaicum, Cirsium heterophyllum, Dryopteris expansa, D. cartusiana) are sometimes found in the community. The moss-lichen layer is characterized by low coverage. Rhodobryum roseum, Rhytidiadelphus triquetrus, Pleurozium schreberi, etc are found here.
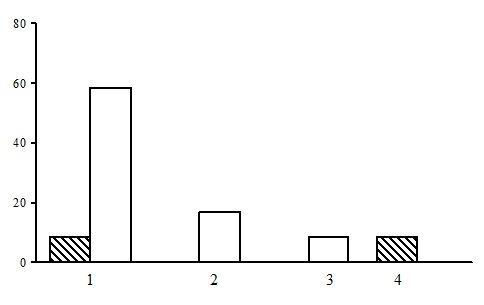
Figure 28. Ratio of PLPs by groups of forest types. Grass and grassmarsh sections. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows groups of forest types (1 – small grass birch/aspen forests, 2 – tall herb spruce forests, 3 – tall herb birch forests, 4 – grassmarsh birch forests),
the Y-axis shows the percentage
Table 29. Some geobotanical parameters of the small grass birch/aspen forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
|
Stand age, yrs |
30.0 |
30.0 |
100.0 |
60.0 |
26.9 |
|
Stand height, m |
9.0 |
9.0 |
25.0 |
18.6 |
5.2 |
|
Tree canopy density (A), % |
80.0 |
33.8 |
85.0 |
68.9 |
16.6 |
| Density of the layer of shrubs and tree understorey (B), % |
14.3 |
2.3 |
38.8 |
15.3 |
12.0 |
| Total projective cover of the grass layer (C), % |
58.8 |
24.9 |
103.2 |
52.7 |
28.9 |
| Total projective cover of the moss-lichen layer (D), % |
9.1 |
4.0 |
31.1 |
11.4 |
9.0 |
| Total projective cover of green mosses in the moss-lichen layer, % |
9.0 |
3.4 |
13.6 |
8.3 |
3.8 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
0.0 |
0.3 |
0.0 |
0.1 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
0.0 |
17.5 |
3.1 |
6.5 |
|
Number of PLPs |
1 |
7 |
|||
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. |
|||||

Figure 29. Small grass birch forest
Four forest types are subdivided within the group of small grass birch/aspen forests: (bunchgrass-bilberry birch forests, forb-bilberry birch forests, horsetail birch forests, and forb-bilberry aspen forests), which are in the majority found in the middle taiga. Forb-bilberry birch forests are predominant (Fig. 30).
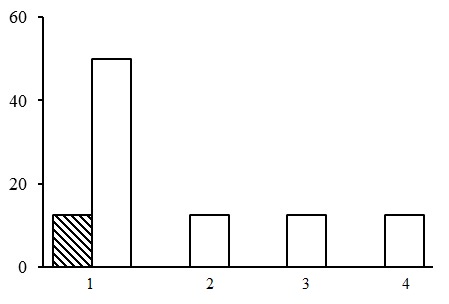
Figure 30. The ratio of forest types in the group of small grass birch/aspen forests. The shaded columns stand for the northern taiga, the unshaded columns – for the middle taiga. The X-axis shows forest types (1 – forb-bilberry birch forests, 2 – bunchgrass-bilberry birch forests, 3 – horsetail birch forests, 4 – forb-bilberry aspen forests), the Y-axis shows the percentage
This group of forest types unites birch forests that appeared on the site of small grass-green moss spruce forests of the middle taiga after logging and fires. These communities grow on drained habitats, and are found on the slopes of moraine hills, on flat sections of dividing ridges and slopes of river terraces on both noncalcareous and carbonate-rich moraine loams.
Table 30. Some geobotanical parameters of the tall-grass spruce forests
|
Parameters |
Forest area |
|||
|
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
|
|
Stand age, yrs |
100.0 |
120.0 |
110.0 |
14.1 |
|
Stand height, m |
25.0 |
26.0 |
25.5 |
0.7 |
|
Tree canopy density (A), % |
45.0 |
67.5 |
56.3 |
15.9 |
| Density of the layer of shrubs and tree understorey (B), % |
7.0 |
15.0 |
11.0 |
5.7 |
| Total projective cover of the grass layer (C), % |
70.2 |
70.8 |
70.5 |
0.4 |
| Total projective cover of the moss-lichen layer (D), % |
27.4 |
31.0 |
29.2 |
2.5 |
| Total projective cover of green mosses in the moss-lichen layer, % |
27.4 |
31.0 |
29.2 |
2.5 |
| Total projective cover of lichens in the moss-lichen layer, % |
0.0 |
0.0 |
0.0 |
0.0 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
0.0 |
0.0 |
0.0 |
|
Number of PLPs |
2 |
|||
| Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. | ||||
Tall herb spruce forests (geobotanical description – see Table 30). The tree layer is formed by the predominant Picea abies with an admixture of Betula pubescens and Populus tremula. Alnus incana, Sorbus aucuparia, Picea abies etc. were found in the layer of shrubs and understorey of trees. The restocking of Picea abies is most common there. The grass-dwarf shrub layer is codominated by Aconitum septentrionale, Vaccinium myrtillus, Equisetum sylvaticum, Oxalis acetosella as well as tall ferns. Some nemoral species are present with low abundance. The moss-lichen layer has low coverage.
Within the group of tall herb and small grass spruce forests one forest type was identified – aconite-tall fern spruce forest.
Tall herb birch forests and grassmarsh birch forests in the studied forests are found within one PLP each (geobotanical description – see Table 31). These forests are represented by communities with the tree layer dominated by Betula pubescens with a small admixture of Picea abies (Fig. 31). The shrubs layer and the understorey are formed by Picea abies, Sorbus aucuparia, etc. The total share of grasses in the grass-dwarf shrub layer exceeds boreal shrubs in terms of coverage. The moss and lichen layer is almost undeveloped.

Figure 31. Tall herb birch forest
High-grass birch forests and grassmarsh birch forests are mainly found in floodplains, drain hollows, in habitats with increased moisture. Such communities are also found on dividing ridges and smooth slopes. Since the understorey of Picea abies is almost always seen in birch forests, such forests can gradually be replaced by spruce forests with an admixture of birch.
Table 31. Some geobotanical parameters of the tall-grass and grassmarsh birch forests
|
Parameters |
Forest type |
|
|
БВ |
ББТ |
|
| Stand age, yrs |
— |
— |
|
Stand height, m |
80.0 |
60.0 |
|
Tree canopy density (A), % |
25.0 |
9.0 |
| Density of the layer of shrubs and tree understorey (B), % |
43.8 |
68.8 |
| Total projective cover of the grass layer (C), % |
4.8 |
1.3 |
| Total projective cover of the moss-lichen layer (D), % |
75.4 |
49.6 |
| Total projective cover of green mosses in the moss-lichen layer, % |
3.9 |
50.6 |
| Total projective cover of lichens in the moss-lichen layer, % |
3.9 |
28.2 |
| Total projective cover of sphagnum mosses in the moss-lichen layer, % |
0.0 |
0.0 |
| Number of PLPs |
1 |
1 |
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. ЕТБрС – grass—cowberry sphagnum spruce forest, БХС – horsetail—sphagnum birch forest. |
||
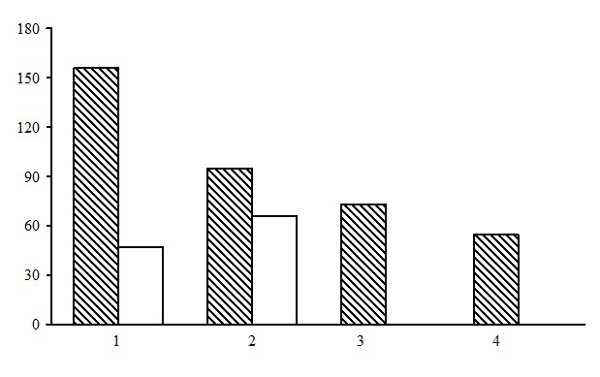
Figure 32. Alpha diversity of forests of the grass and grass-marsh sections. The shaded columns stand for the total number of species in the forest type group (species richness); the unshaded columns – for the average number of species per PLP (species density). The X-axis shows groups of forest types (1 – small grass birch/aspen forests, 2 – tall herb spruce forests, 3 – tall herb birch forests 4 – grassmarsh birch forests), the Y-axis – the number of plant species (vascular plants, mosses, lichens)
The floristic diversity of forests of the grass and grass-marsh sections was estimated using alpha and beta diversity indicators.
Evaluation of alpha diversity. Species density in different groups of forest types of the grass and grass-marsh sections varies: from 47 to 66 species (Fig. 32). At the same time, the majority of PLPs had 40 to 60 plant species (Tables 32–34). Tall-grass spruce forests and small-grass birch forests have the highest values of floristic diversity. The tall-grass spruce forests have lower species richness as compared to small-grass birch forests due to the sample size: birch forests were found on eight PLPs, whereas spruce forests – only on two.
Evaluation of beta diversity. Beta diversity (Tables 35 and 36) within the grass and grass-marsh sections is assessed using the value of Jaccard index and Whittaker’s index.
Pair-wise comparison of groups of forest types of the grass and grass-marsh sections shows that Jaccard index of similarity varies from 0.2 to 0.6. Tall herb birch and spruce forests demonstrated the highest similarity in floristic composition (0.6); while the lowest similarity was demonstrated by grassmarsh birch forests and small grass birch forests (0.2). The value of Whittaker’s index (4.3) for the grass and grass-marsh sections is higher than for groups of the forest types within the section (1.5–3.4).
Table 32. Some parameters of the floristic diversity in the small grass birch/aspen forests
|
Parameters |
Forest area |
||||
|
Northern taiga |
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
||
| Number of species per PLP (vascular plants, mosses and lichens) |
37.0 |
29.0 |
70.0 |
48.4 |
17.3 |
|
Number of vascular plants per PLP |
28.0 |
18.0 |
47.0 |
32.6 |
12.5 |
| Average number of vascular plant species per a PLP plot |
16.8 |
9.3 |
26.0 |
18.0 |
6.1 |
|
Number of mosses and lichens species per PLP |
9.0 |
7.0 |
24.0 |
15.9 |
7.0 |
| Average number of moss and lichen species per a PLP plot |
5.8 |
2.0 |
11.5 |
6.9 |
3.1 |
|
Number of PLPs |
1 |
7 |
|||
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. |
|||||
Table 33. Some parameters of the floristic diversity of the tall herb spruce forests
|
Parameters |
Forest area |
|||
|
Middle taiga |
||||
|
min |
max |
x |
st. dev. |
|
|
Number of species per PLP (vascular plants, mosses and lichens) |
57.0 |
68.0 |
62.5 |
7.8 |
|
Number of vascular plants per PLP |
42.0 |
46.0 |
44.0 |
2.8 |
| Average number of vascular plant species per a PLP plot |
23.5 |
29.0 |
26.3 |
3.9 |
|
Number of mosses and lichens species per PLP |
11.0 |
26.0 |
18.5 |
10.6 |
| Average number of moss and lichen species per a PLP plot |
7.8 |
9.5 |
8.6 |
1.2 |
|
Number of PLPs |
2 |
|||
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. |
||||
Table 34. Some parameters of the floristic diversity of the tall herb and grassmarsh birch forests
|
Parameters |
Forest type |
|
|
БВ |
ББТ |
|
| Number of species per PLP (vascular plants, mosses and lichens) |
69.0 |
51.0 |
|
Number of vascular plants per PLP |
49.0 |
25.0 |
|
Average number of vascular plant species per PLP plot |
24.0 |
13.0 |
|
Number of mosses and lichens species per PLP |
20.0 |
26.0 |
| Average number of moss and lichen species per PLP plot |
8.0 |
15.0 |
|
Number of PLPs |
1 |
1 |
|
Comment. PLP – permanent lookout point, min – minimum value, max – maximum value, x – average value, st. dev. – standard deviation. ЕТБрС – grass—cowberry sphagnum spruce forest, БХС – horsetail—sphagnum birch forest. |
||
Table 35. Jaccard index values for groups of the forest types of the grass and grass-marsh sections
|
Forest type group |
Б/ОМ |
ЕВ |
БВ |
ББТ |
|
Б/ОМ |
1 |
0.3 |
0.4 |
0.2 |
|
ЕВ |
— |
1 |
0.6 |
0.3 |
|
БВ |
— |
— |
1 |
0.2 |
|
ББТ |
— |
— |
— |
1 |
| Comment. Б/ОМ – small grass birch/aspen forests, ББТ – grassmarsh birch forests,
БВ – tall herb birch forests, ЕВ – tall herb spruce forests |
||||
Table 36. Beta diversity values of the forests of the grass and grass-marsh sections
|
Forest type group |
Б/ОМ |
ЕВ |
БВ |
ББТ |
For the section |
|
Total number of species |
156.0 |
95.0 |
73.0 |
55.0 |
224.0 |
|
Average species density |
47.0 |
66.0 |
— |
— |
53.0 |
|
Whittaker index (W) |
3.4 |
1.5 |
— |
— |
4.3 |
| Comment. Б/ОМ – small grass birch/aspen forests, ББТ – grassmarsh birch forests, БВ – tall herb birch forests, EB – tall herb spruce forests | |||||
Ecological-coenotic structure of forests. The results of the floristic list–based calculation of the ecological-coenotic structure of species diversity have shown that forests of the grass and grass-marsh sections have a fairly diverse set of ecological-coenotic plant groups (Fig. 33). These forests are dominated by boreal forest species of all life forms: trees, shrubs, dwarf shrubs and grasses. The share of nemoral and wetland types is high. Nitrophilous plants were found. The share of meadow and forest-edge plants is insignificant, which is typical for the taiga forests.
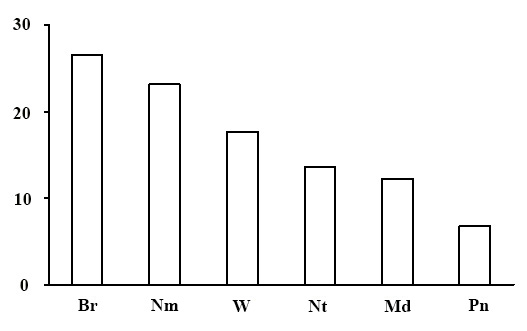
Figure 33. Ecological-coenotic structure of forests of the grass and grass-marsh sections. The X-axis shows ecological-coenotic plant groups (Br – boreal, Nm – nemoral, Pn – pine-forest, Md – meadow and meadow-forest edge, Nt – nitrophilous, W – wetland), the Y-axis shows the percentage
The communities belonging to the grass and grass-marsh sections are not often found in the studied forests and are mainly represented by the birch forests of the middle taiga. The birch forests of the grass section are the forests that have replaced small grass-green moss spruce forests after felling or fires (Morozova et al., 2008). Birch forests of the grassmarsh section are mainly found in floodplains and wetland terrace-near depressions. Since the understorey of spruce is almost always present in birch forests, such forests may gradually be replaced by spruce forests with an admixture of birch. Tall herb forests – birch and spruce forests – are the richest in floristic and structural diversity. Such communities are similar to each other.
CONCLUSION
The classification of forests was carried out for the territory of Karelia and the Karelian Isthmus using the field guide of forest types of the European Russia developed by L. B. Zaugolnova and V. B. Martynenko. The studied forests were divided into five main sections: lichenous, green moss, sphagnous, grass and grassmarsh. The most common sections were the green moss and the sphagnous ones. They include five groups of forest types each. The least diverse is the lichenous section: only two groups of forest types are found there. Individual forest types were identified in each forest types group. The most common types of forest were bilberry-green moss pine and spruce forests.
The majority of the studied forests (60%) was located in the middle taiga and the smaller part (40%) – in the northern taiga. Main forest-forming species forming the trees layer are: common pine (Pinus sylvestris), Norway spruce (Picea abies), common birch (Betula pendula) and pubescent birch (B. pubescens). The shrubs layer and the understorey consist of the young generation of the above-named trees and adult boreal shrubs. Dwarf shrubs, evergreen grasses and boreal small grasses are common in the grass-dwarf shrub layer; forests with the predominance of boreal tall herb and large ferns are rare. The forests of the northern taiga differ from the forests of the middle taiga: the tree canopy is less dense and heathberry (Empetrum sp.) is present in different forest types. Lichens, green and sphagnum mosses can be combined in different ways in the moss-lichen layer. At the same time, both literature data and geobotanical descriptions show that the majority of the studied forests are secondary forests of post-felling or post-fire origin. It is proven by the age of the main forest-forming species (they are young, mostly younger than 100 years); by the closed stands, which are usually formed after major disturbances (felling, fire); by a high share of pioneer species of trees (common pine, common birch and pubescent birch) that are the first to settle on open treeless spaces.
The highest species richness is shown by the forests of the green moss section, whereas the highest species density is shown by the forests of the grass section. High species richness is due to the significant occurrence of the forests of this section in the study area and therefore a large number of the established PLPs. High species density is determined by a large number of species on the grass section PLPs. The lowest values of species richness and density are found in the lichenous section. At the same time, all the studied forests have a fairly diverse set of ecological-coenotic plant groups. Boreal forest species of all life forms are dominating here: trees, shrubs, dwarf shrubs and grasses, which is typical for taiga forests.
Acknowledgements
The study was carried out as part of the line of study No. 0110-2018-0004 «Conservation and regeneration of ecological functions of forest soils», state assignment «Methodical approaches to the assessment of the structural organization and functioning of forest ecosystems»№ АААА-А18-118052400130-7, results of the study were processed by supported Russian Science Foundation (project 16-17-10284).
The author would like to express gratitude to T.Yu. Braslavskaya, N.V. Genikova, E.E. Kostina, and N.V. Ivanova for geobotanical descriptions made.
REFERENCES
Aleksandrova V.D. Klassifikaciya rastitel’nosti: obzor principov klassifikacii i klassifikacionnyh sistem v raznyh geobotanicheskih shkolah (Classification of vegetation: principles and classification systems of various phytocoenological scools), Leningrad: Nauka, 1969, 275 p.
Braslavskaya T.Yu., Zaugolnova L.B. Klassifikaciya i tipologiya lesnoj rastitel’nosti kak instrument issledovaniya lesnogo pokrova (Classification and typology of forest vegetation as a tool for the study of forest cover), In: Metodicheskie podkhody k ekologicheskoj otsenke lesnogo pokrova v bassejne maloj reki (Methodical approaches to the ecological assessment of forest cover in the small river basin), Moscow: Tovarishchestvo nauchnykh izdanij CMC, 2010, рp. 56-71.
Braun-Blanquet J. Pflanzensoziologie, Wien-N.Y: Springer, 1964, 565 s.
Bulokhov A.D., Solomeshch A.I. Ekologo-floristicheskaya klassifikatsiya lesov Yuzhnogo Nechernozemya (Ecological and floristic classification of forests of the Southern Non-Black Earth Region), Bryansk: Izd-vo Bryanskogo gos. ped. un-ta, 2003, 359 p.
Cherepanov S.K. Sosudistye rasteniya Rossii i sopredel’nyh gosudarstv (Vascular plants of Russia and neighboring states), St. Petersburg: “Mir i Sem’ya-95”, 1995, 992 p.
Geobotanicheskoe rajonirovanie Nechernozemya evropejskoj chasti RSFSR (Geobotanical zoning of the Black Earth Region of the European Part of the RSFSR), Ed. ed. V.D. Aleksandrova, T.K. Yurkovskaya, Leningrad: Nauka, 1989, 64 p.
Korchagin A.A. Rastitelnost severnoj poloviny Pechoro-Ilychskogo zapovednika (Vegetation of the northern half of the Pechora-Ilych reserve), Trudy Pechoro-Ilychskogo zapovednika, 1940, Issue 2, 416 p.
Lukina N.V., Orlova M.A., Gornov A.V., Kryshen’ A.M., Kuznecov P.V., Knyazeva S.V., Smirnov V.E., Bahmet O.N., Edlina S.P., Ershov V.V., Zukert N.V., Isaeva L.G. Ocenka kriteriev ustojchivogo upravleniya lesami s ispol’zovaniem indikatorov mezhdunarodnoj programmy ICP Forests (Evaluation of criteria for sustainable forest management using indicators of the international program ICP Forests), Lesovedenie. 201, No 5, pp. 62-75.
Martynenko V.A. Temnohvojnye lesa (Dark coniferous forests), In: Lesa Respubliki Komi (Forests of the Komi Republic). Moscow: Dizajn. Informaciya. Kartografiya, 1999, рp. 133-184.
Metodika monitoringa lesov po mezhdunarodnoj programme ICP Forests (Forest monitoring method of the international program ICP Forests), Moscow, 2008, 46 p.
Mirkin B.M., Naumova L.G. Nauka o rastitel’nosti (istoriya i sovremennoe sostoyanie osnovnyh koncepcij) (The science of vegetation (history and current state of the basic concepts)) Ufa: Gilem, 1998,413 p.
Mirkin B.M., Rozenberg L.G., Naumova L.G. Slovar’ ponyatij i terminov sovremennoj fitocenologii (Dictionary of concepts and terms of modern phytocenology), Moscow, 1989, 224 p.
Morozova O.V., Zaugol‘nova L.B., Isaeva L.G., Kostina V.A. Klassifikaciya boreal‘nyh lesov severa Evropejskoj Rossii. Oligotrofnye hvojnye lesa (Classification of boreal forests of northern European Russia. Oligotrophic coniferous forests), Rastitel’nost’ Rossii, 2008, No 13, pp. 61—81.
Nicenko A.A. Ob izuchenii ehkologicheskoj struktury rastitel’nogo pokrova (On the study of the ecological structure of vegetation), Botanicheskii zhurnal, 1969, Vol. 54, No 7, pp. 1002-1014.
Volkov A.D. Tipy lesa Karelii (Types of Karelia forest), Petrozavodsk: Karelskii NTs RAN, 2008, 180 p.
Vostochnoevropejskie lesa: istoriya v golotsene i sovremennost’ (Eastern European forests: history in the Holocene and modernity), Book 1. Moscow: Nauka, 2004, 479 p.
Yurkovskaya T.K., Elina G.A. Vosstanovlennaya rastitel’nost’ Karelii na geobotanicheskoj i paleokartah (Restored vegetation of Karelia on geobotanical and paleo-maps), Petrozavodsk: Karelskii NTs RAN, 2009, 136 p.
Zaugolnova L.B., Martynenko V.B. Opredelitel’ tipov lesa Evropejskoj Rossii (Determinant of forest types in European Russia), Available at: http://cepl.rssi.ru/bio/forest/index.htm (2018, 01 November).
Zaugolnova L.B., Morozova O.V. Rasprostranenie i klassifikatsiya borealnykh lesov (Distribution and classification of boreal forests), In: Vostochnoevropejskie lesa: istoriya v golotsene i sovremennost’ (Eastern European forests: history in the Holocene and modernity). Book 2. Moscow: Nauka, 2004, рp. 295-330.
Zaugolnova L.B., Morozova O.V. Tipologiya i klassifikatsiya lesov Evropejskoj Rossii metodicheskie podkhody i vozmozhnosti realizatsii (Typology and classification of forests in European Russia: methodical approaches and opportunities for implementation), Lesovedenie, 2006, No 1, рp. 34-48.
Zozulin G.M. Istoricheskie svity rastitelnosti Evropejskoj chasti SSSR (Historical formation of vegetation of the European part of the USSR), Botanicheskii zhurnal, 1973, Vol. 58, No 8, рp. 1081-1092.
Reviewer: PhD in biology Panasenko N.N.





