- DOI 10.31509/2658-607x-2020-3-4-1-25
- УДК 582.675.1, 582.573.36, 582.579.2: 581.522
Влияние одиночных деревьев на флористическое разнообразие и популяционную структуру редких видов растений остепненных лугов
![]()
© 2020 г. Е.В. Ручинская*, А.В. Горнов
Центр по проблемам экологии и продуктивности лесов РАН
Россия, 117997 Москва, ул. Профсоюзная, 84/32, стр. 14
*E-mail: elena.ruchinskaya@mail.com
Поступила в редакцию: 01.11.2020
Принята к печати: 14.12.2020
В зоне широколиственных лесов Европейской России сохранились остепненные луга, которые обладают богатым флористическим составом и вносят значительный вклад в биологическое разнообразие территорий. Один из лесных регионов России, где встречаются такие луга, – Брянская область. Здесь уцелели остепненные луга с высоким флористическим разнообразием и большим числом редких видов растений. На эти луга постоянно внедряются деревья из окружающих лесных участков. Большая часть молодых деревьев погибает от регулярных палов и хозяйственной деятельности. Однако некоторые особи выживают и переходят в генеративное состояние, в котором они относительно устойчивы к низовым пожарам. Влияние одиночных деревьев на флористическое разнообразие остепненных лугов изучено на двух уровнях организации живых систем – ценотическом и популяционном. На ценотическом уровне исследованы полидоминантные остепненные луга и полидоминантные остепненные луга с одиночными генеративными деревьями, на популяционном уровне – ценопопуляции Iris aphylla, Anemone sylvestris и Anthericum ramosum. При сборе материала использованы разные методы: геоботанические, демографические, измерения факторов среды (освещенность, крутизна склонов и периодичность палов). Установлено, что полидоминантные остепненные луга сохранились в средней части крутых склонов, где затруднены выпас и сенокошение, а также нечасто случаются палы травы. Эти сообщества обладают высоким флористическим разнообразием и устойчивыми ценопопуляциями модельных видов. Онтогенетические спектры Anemone sylvestris, Anthericum ramosum и Iris aphylla относятся к полночленному левостороннему типу с максимальной численностью особей. Одиночные деревья (Quercus robur, Tilia cordata) неоднозначно влияют растительность полидоминантных остепненных лугов. С одной стороны, с появлением деревьев возрастает видовое разнообразие сообществ. Это связано с тем, что деревья – удобные места отдыха и укрытия для птиц, которые разносят диаспоры растений. С другой стороны, взрослые деревья затеняют травяной покров. Это приводит к сокращению покрытия и встречаемости степных и сухолуговых видов, а также влияет на их популяционную структуру. Онтогенетический спектр Anemone sylvestris остается полночленным, Iris aphylla становится неполночленным, а Anthericum ramosum –незавершенным.
Ключевые слова: остепненные луга, одиночные деревья, флористическое разнообразие, ценопопуляция, онтогенетический спектр, Anemone sylvestris, Anthericum ramosum, Iris aphylla
В зоне широколиственных лесов Европейской России сохранились остепненные луга (Булохов, 1977, 2001; Босек, 1980; Скворцов, 1982; Аверинова, 2010; Семенищенков, 2010, 2012; Евстигнеев и др., 2011; Панасенко и др., 2013, 2015 и др.). Эти сообщества, как правило, обладают богатым флористическим составом и вносят значительный вклад в биологическое разнообразие территорий. Однако из-за хозяйственной деятельности человека и палов травы такие ценозы находятся под угрозой исчезновения (Зеленая…, 2012; Евстигнеев и др., 2018а; Ручинская, 2019). На остепненные луга, сохранившиеся в зоне широколиственных лесов, постоянно внедряются древесные растения из окружающих лесных участков. Большая часть молодых деревьев погибает от регулярных палов и хозяйственной деятельности: сенокошения, выпаса и др. Некоторые особи выживают и переходят в генеративное состояние (Евстигнеев и др., 2018а). Взрослые деревья относительно устойчивы к низовым пожарам: их почки возобновления находятся высоко, а толстая корка ствола защищает камбий (Серебряков, 1962). Одиночные деревья влияют на условия произрастания других растений на лугах. Известно, что в фитогенных полях деревьев существенно изменяются освещенность, температура и влажность воздуха, температура и влажность почвы, количество осадков, проникающих сквозь крону, качество опада, концентрация элементов питания и другие почвенные характеристики (Уранов, 1965; Самойлов, 1983; Никонов и др., 2002; Ипатов, 2007; Журавлева и др., 2012; Орлова и др., 2016). Кроме того, одиночные деревья привлекают животных разных экологических групп: почвенных беспозвоночных, мышевидных грызунов, птиц и др. (Manning et al., 2006; Prevedello et al., 2018). С одной стороны, они влияют на условия произрастания растений, а с другой – участвуют в создании как внутриценотических, так и межценотических потоков диаспор. В связи с этим в работе поставлена цель – рассмотреть влияние одиночных деревьев на флористический состав и состояние ценопопуляций некоторых редких видов растений остепненных лугов.
Материал и методы
Исследования проведены на юго-востоке Брянской области в пределах памятника природы «Меловицкие склоны» (рис. 1). Участок расположен в Комаричско-Севском физико-географическом районе. Он представляет собой возвышенные лессовые равнины с оврагами, балками, склонами и выходами карбонатных пород на западных отрогах Среднерусской возвышенности. В ботанико-географическом плане территория относится к Восточноевропейской провинции Европейской широколиственно-лесной области (Растительность…, 1980). Климат Комаричско-Севского района умеренно континентальный. Среднегодовая температура 5.4οС. Продолжительность теплого времени года с температурой выше 0°С − 228 суток, вегетационного периода с температурой выше + 5°C – 188 суток. Среднегодовое количество осадков – 613 мм, среднее количество осадков в теплый период года – 342 мм (Природное…, 1975).
Исследования проводили на двух уровнях организации живых систем – ценотическом и популяционном. На ценотическом уровне изучали полидоминантные остепненные луга и полидоминантные остепненные луга с одиночными генеративными деревьями. На популяционном уровне объектами исследования стали ценопопуляции модельных видов растений: Iris aphylla L. (касатик, ирис безлистный), Anemone sylvestris L. (ветреница лесная) и Anthericum ramosum L. (венечник ветвистый). Iris aphylla – короткокорневищное розеточное весеннецветущее летнезеленое растение (рис. 2, А). Геофит. Anemone sylvestris – многолетнее травянистое весеннецветущее летнезеленое короткокорневищное растение (рис. 2, Б). Гемикриптофит и геофит. Anthericum ramosum – многолетнее травянистое летнезеленое летнецветущее короткокорневищное растение (рис. 3). Гемикриптофит и геофит. Выбор этих видов обусловлен тем, что они редки, исчезают и занесены в Красные книги многих регионов (Красная…, 2002, 2004, 2015, 2016 и др.). Кроме того, Iris aphylla занесен в Красную книгу России (2008).

Рисунок 1. Расположение памятника природы «Меловицкие склоны». Зеленая линия – границы объекта исследований. Фоновое спутниковое изображение – Microsoft Bing Maps
В работе использованы следующие методы исследования: геоботанические, демографические, измерения факторов среды (освещенности, крутизны склонов и периодичности палов) и статистические. Геоботанические описания сделаны на площадках по 100 м2 в 11-кратной повторности в каждом варианте сообществ. На каждой площадке составляли полный флористический список. Участие видов оценено в баллах по шкале обилия-покрытия Ж. Браун-Бланке (Миркин и др., 1989). Для оценки видового разнообразия сообществ использовали видовое богатство и видовую насыщенность. Видовое богатство – суммарное число видов в сообществе, которое получено на основе 11 геоботанических описаний. Видовая насыщенность – среднее число видов на единицу площади. Названия сосудистых растений даны по данным международной базы The Plant List (http://www.theplantlist.org/). Для анализа геоботанических описаний сообществ применили ординацию методом бестрендового анализа соответствий – Detrended Correspondence Analysis (DCA). Этот метод эффективно работает с гетерогенными данными геоботанических описаний (Джонгман и др., 1999). Расчеты проводили в программе PC-ORD. В основу демографических исследований легла периодизация онтогенеза, предложенная Т.А. Работновым (1950) и дополненная А.А. Урановым (1975) и его учениками (Ценопопуляции…, 1988). Онтогенез делится на этапы, отличающиеся морфологически и функционально. Особи, относящиеся к одному онтогенетическому состоянию, объединяются в одну группу: j – ювенильные, im – имматурные, v – виргинильные, g1 – генеративные молодые, g2 – генеративные средневозрастные, g3 – генеративные старые, ss – субсенильные, s – сенильные. Онтогенетические состояния модельных видов определяли на основе публикаций (Евстигнеев и др., 2018; Ручинская, 2019). Состояние ценопопуляций оценивали с помощью численности, плотности и типа онтогенетического спектра. Численность – число особей на исследуемой территории (Чернова, Былова, 2007). Плотность ценопопуляции – среднее число особей на единицу площади (Одум, 1986; Ценопопуляции…, 1988). Тип онтогенетического спектра называли по классификации, предложенной ранее (Заугольнова, 1994). На лугах и под деревьями ежечасно определяли освещенность с помощью люксметра в безоблачный июньский день с 10 до 18 ч. Люксы переводили в проценты от полной освещенности, которую определяли на открытом месте. Крутизну склона измеряли с помощью дальномера-угломера Nikon Forestry Pro. Периодичность палов определяли по возрасту побегов формирования у кустарников (Frangula alnus Mill., Corylus avellana L.). Эти побеги появляются из спящих почек, расположенных в базальной части кустарника, прежние надземные оси которого погибли после повреждения огнем (рис. 4).
Результаты и обсуждение
Полидоминантные остепненные луга сохранились в средней части крутых склонов, где затруднены выпас и сенокошение (рис. 5). Палы случаются преимущественно один раз в два года. Они ограничивают внедрение древесных растений – молодые особи деревьев наиболее уязвимы. Например, проростки и ювенильные растения дуба часто погибают при палах травы (Комаров, 1951). В результате формируются полидоминантные сообщества с высоким видовым разнообразием (табл. 1; приложение). Уникальность ценозам придают виды, которые свойственны степным сообществам: Ajuga genevensis L., Anemone sylvestris, Aster amellus L., Astragalus cicer L., Campanula sibirica L., Prunus cerasus L., Galium tinctorium L., G. verum L. и др. В эколого-ценотической структуре преобладают растения сухолуговой группы, в которую входят и перечисленные степные. Нередко встречаются влажно-луговые (Festuca pratensis Huds., Hypericum maculatum Crantz, Succisa pratensis Moench, Thalictrum lucidum L. и др.), неморально-опушечные (Brachypodium pinnatum (L.) Beauv., Peucedanum cervaria (L.) Cusson ex Lapeyr., Laserpitium latifolium L., Lathyrus pisiformis L., L. sylvestris L., Pyrethrum corymbosum (L.) Scop.) и черноольхово-опушечные (Rubus caesius L. и Valeriana officinalis L.) растения. Небольшое участие характерно для лесных видов: неморальных – Convallaria majalis L., Corylus avellana, Quercus robur L., Viola mirabilis L., бореальных – Frangula alnus, боровых – Pteridium aquilinum, Solidago virgaurea L. и Viola collina Besser. Диаспоры лесных и опушечных видов заносятся сюда животными и ветром из соседнего сосняка, а влажно-луговых и черноольховых – из пойменных сообществ, которые примыкают к склону.
Таблица 1. Характеристики сообществ на остепненных склонах.
Памятник природы «Меловицкие склоны»
| Показатели | Сообщества | |
| 1 | 2 | |
| Угол склона | ||
| Угол склона, M ± σ | 37 ± 2.4 | 31 ± 2.4 |
| Диапазон угла склона | 33–41 | 28–37 |
| Число измерений | 21 | 33 |
| Пожары | ||
| Частота пожаров, M ± σ | 2.3 ± 1.2 | 2.2 ± 1.0 |
| Число измерений | 52 | 33 |
| Характеристика разнообразия видов сосудистых растений | ||
| Среднее число видов на 100 м2, M ± mM | 51 ± 1.2 | 59 ± 1.2 |
| Диапазон числа видов на 100 м2 | 44–56 | 52–66 |
| Число видов на 11 площадках по 100 м2 | 98 | 107 |
| Число и доля (%) видов разных эколого-ценотических групп | ||
| Сухолуговая | 77 (78.6) | 79 (73.8) |
| Влажно-луговая | 5 (5.1) | 7 (6.5) |
| Неморальная лесная | 4 (4.1) | 8 (7.5) |
| Неморальная опушечная | 6 (6.1) | 6 (5.6) |
| Боровая | 3 (3.1) | 3 (2.8) |
| Бореальная лесная | 1 (1.0) | 1 (1.0) |
| Черноольховая опушечная | 2 (2.0) | 3 (2.8) |
Примечание. M – среднее арифметическое, σ – стандартное отклонение. Сообщества: 1 – полидоминантные остепненные луга, 2 – полидоминантные остепненные луга с одиночными генеративными деревьями
Iris aphylla – один из преобладающих видов в травостое полидоминантных остепненных лугов «Меловицких склонов». Плотность ценопопуляции касатика составляет 82 особи 1 м2. Онтогенетический спектр – полночленный одновершинный с максимумом на v и gн особях (рис. 6, 1а). Касатик хорошо приспособлен к условиям высокой освещенности открытых пространств, благодаря строению своих листьев: они уплощены с боков и ориентированы вертикально (Евстигнеев и др., 2018б). Семенному возобновлению ириса способствует деятельность животных – муравьев и мышевидных грызунов, которые населяют склоны и создают нарушения. Эти микросайты отличаются разреженным травяным покровом, разрыхленным субстратом, повышенными аэрацией и температурой почвы, значительной микробиологической активностью (Зрянин, 2003; Dauber, Wolters, 2000; Kostrakiewicz, 2004 и др.). Например, на выбросе мышевидного грызуна площадью 0.03 м2 встречен популяционный локус, состоящий из 10 ювенильных особей. Распространению диаспор касатика способствуют муравьи (рис. 7). Р.Е. Левина (1957) утверждает, что свежие семена привлекают этих животных сладкой клейкой жидкостью, которые содержатся в оболочке. Наши наблюдения показали, что муравьи также распространяют и сухие семена (Евстигнеев и др., 2018б).

Рисунок 2. Модельные виды растений на остепненных лугах памятника природы «Меловицкие склоны»: А – Iris aphylla, Б – Anemone sylvestris. Фото – Горнов А.В.

Рисунок 3. Модельные виды растений на остепненных лугах памятника природы «Меловицкие склоны»: А, Б – Anthericum ramosum. Фото – Ручинская Е.В.

Рисунок 4. Порослевые побеги обгоревшей вишни кустарниковой (Prunus cerasus).
А – общий вид кустарника, Б – основание кустарника. 1 – пень от обгоревшего многолетнего побега, 2 – мертвый обгоревший двулетний порослевой побег, 3 – живой однолетний порослевой побег, который проснулся из спящей почки после низового пожара весной (по: Ручинская, 2019).

Рисунок 5. Полидоминантные остепненные луга на территории памятника природы «Меловицкие склоны». Фото – Горнов А.В.

Рисунок 6. Онтогенетический спектр ценопопуляций модельных видов растений на остепненных лугах: 1 – Iris aphylla, 2 – Anthericum ramosum, 3 – Anemone sylvestris. По оси абсцисс – онтогенетические состояния, по оси ординат – доля особей, %. В кружке – плотность ценопопуляции (число особей на 1 м2). Сообщества: а – полидоминантные остепненные луга, б – полидоминантные остепненные луга с одиночными генеративными деревьями. Онтогенетические состояния особей: j – ювенильное, im – имматурное, v – виргинильное, g1 – молодое генеративное, g2 – зрелое генеративное, g3 – старое генеративное, ss – субсенильное, s – сенильное

Рисунок 7. Перемещение свежих семян Iris aphylla рыжим лесным муравьем (Formica rufa). Фото – Ручинская Е.В.
Anthericum ramosum – содоминант в травостое остепненных лугов. Плотность ценопопуляции венечника составляет 56 особей на 1 м2. Онтогенетический спектр – полночленный левосторонний одновершинный с максимумом на v и gн особях (рис. 6, 2а). Формирование максимума на v и gн особях определяется: 1) короткой длительностью j и im состояний; 2) пополнением за счет особей, отдыхающих от цветения; 3) пополнением v особями вегетативного происхождения, которые образуются в результате дезинтеграции g2 растений.
Anemone sylvestris может быть как ассектатором, так и содоминантом в травостое остепненных лугов. Плотность ценопопуляции ветреницы составляет – 75 особей на 1 м2. Онтогенетический спектр полночленный левосторонний с максимумом на im растениях (рис. 6, 3а), плотность которых составляет 27 особей на 1 м2. В исследуемом сообществе не отмечены особи ветреницы семенного происхождения. Это позволяет назвать спектр вегетативно-полночленным. Высокая плотность ценопопуляции и полночленный онтогенетический спектр определяются особенностью биологии ветреницы. Большое число растений прегенеративного периода обусловлено способностью ветреницы к вегетативному размножению с глубоким омоложением (рис. 8). Вегетативные особи развиваются из почек, появляющихся на горизонтальных придаточных корнях (Старостенкова, 1986; Барыкина, Потапова, 1994). Начало вегетации ранней весной, до того как поднимется травостой, способствует накоплению достаточного количества пластических веществ, которые необходимы особям для формирования генеративных органов. Стоит отметить, что отсутствие семенных особей говорит о затрудненных условиях для ценопопуляции. Скорее всего, это связано с распространением огня на склонах – он уничтожает молодые семенные особи ветреницы.
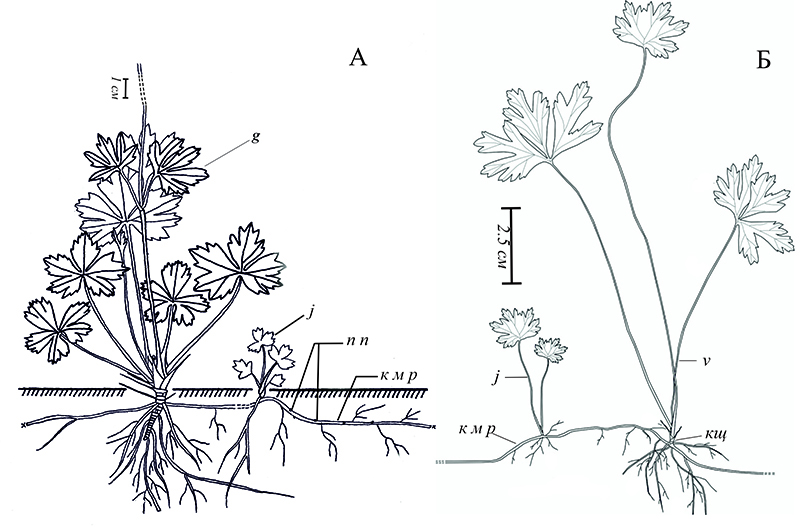
Рисунок 8. Вегетативное возобновление Anemone sylvestris: А – генеративная особь с корневым отпрыском и придаточными почками на корне (по: Барыкина, Потапова, 1994, с дополнением), Б – ювенильная и виргинильная особи корнеотпрыскового происхождения (по: Горнов и др., 2013). j – ювенильная особь, g – генеративная особь, v – виргинильная особь, к м р – корень материнского растения, кщ – корневище, п п – придаточная почка
Полидоминантные остепненные луга с одиночными генеративными деревьями. На склонах встречаются одиночные генеративные деревья Quercus robur и Tilia cordata (рис. 9), которые в виргинильном периоде избежали гибели от огня. Взрослые особи дуба и липы относительно устойчивы к низовым пожарам: их почки возобновления находятся высоко, а толстая корка ствола защищает камбий (Серебряков, 1962). Крутизна склона и частота палов сходна с полидоминантными остепненными лугами. При этом ординация геоботанических описаний четко разделила полидоминантные остепненные луга и полидоминантные остепненные луга с одиночными деревьями на группы (рис. 10). Сообщества отличаются максимальными значениями видового богатства и видовой насыщенности (табл. 1; приложение). Высокое видовое разнообразие определяется несколькими причинами. Во-первых, в прошлом сообщества не подвергались активному выпасу и сенокошению, поскольку также размещены на крутых частях склонов. Во-вторых, одиночные деревья – удобные места отдыха и укрытия для многих животных, в том числе и птиц. Они, как известно, разносят семена луговых и лесных растений (Manning et al., 2006; Prevedello et al., 2018). В результате сообщества с отдельно стоящими деревьями характеризуются более высоким видовым богатством, чем полидоминантные остепненные луга. В эколого-ценотической структуре сообщества преобладают сухолуговые и степные растения. Под кронами одиночных деревьев освещенность снижается до 60% от полной. Притенение сокращает покрытие светолюбивых сухолуговых и степных растений. Однако число видов этой группы увеличивается. Появляются Allium oleraceum L., Artemisia absinthium L., Carex montana L., Cirsium decussatum Janka, Fallopia convolvulus (L.) Á. Löve, Filipendula vulgaris Moench, Hypericum perforatum L., Silene vulgaris (Moench) Garcke, S. viscaria (L.) Jess., Stachys officinalis (L.) Trevis., Veronica spuria L. и др. Кроме того расширяется видовой состав и других эколого-ценотических групп: среди влажно-луговых присутствуют Carex hirta L., C. lachenalii Schkuhr, Galeopsis bifida Boenn; группу неморальных растений пополнили Euonymus europaeus L., Lathyrus niger (L.) Bernh. и Pyrus communis L., а опушечно-черноольховых – Galium aparine L. Скорее всего, это связано с деятельностью животных, в первую очередь птиц, которые прилетая на одиночные деревья, занесли диаспоры перечисленных растений. Известно, что птицы активно разносят семена многих видов растений (Левина, 1957; Cramp, 1998 и др.). Таким образом, благодаря одиночным деревьям поддерживается полидоминантный состав сообщества с максимальным видовым разнообразием. Однако затенение сказывается на состоянии ценопопуляций модельных видов растений.

Рисунок 9. Полидоминантные остепненные луга с одиночными деревьями: А – общий вид склона, Б – крона Tilia cordata сверху. Фото – Ситников А.Ю.
Iris aphylla теряет свои позиции в травостое полидоминантных остепненных лугов с одиночными генеративными деревьями. Плотность ценопопуляции составляет 50 особей на 1 м2. Это почти в два раза ниже, чем в полидоминантных остепненных лугах. Снижение плотности определяется тем, что из-за небольшого светового довольствия у ириса формируется очень мало плодоносящих особей, которые могут сформировать жизнеспособные семена. Это приводит к падению численности молодых семенных растений в ценопопуляции в четыре раза. Прижившиеся особи отличаются увеличенной площадью листовой пластинки (табл. 2). Это является адаптивным приспособлением, благодаря которому растения улавливают больше рассеянного света. Однако из-за нехватки светового довольствия большинство особей ириса развиваются только до v-онтогенетического состояния, а затем – погибают. В результате формируется прерывистый онтогенетический спектр с максимумом на v особях (рис. 6, 1б). Если со временем число деревьев на склоне увеличится, и они сформируют сомкнутый участок леса, то ценопопуляция касатика исчезнет.
Таблица 2. Длина и ширина листьев Iris aphylla на свету (1) и в тени (2)
| Признак | N | M ± mM | σ | U | ||||
| 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | |
| Длина листа, см | 22 | 26 | 31.4 ± 2.22 | 60.6 ± 1.70 | 10.4 | 8.7 | 6 (p = 0.000000) | |
| Ширина листа, см | 22 | 26 | 1.6 ± 0.09 | 2.3 ± 0.08 | 0.4 | 0.4 | 56 (p = 0.000002) | |
Примечание. Сообщества: 1 – полидоминантные остепненные луга, 2 – полидоминантные остепненные луга с одиночными генеративными деревьями. N – число измерений, M – среднее арифметическое, mM – ошибка среднего арифметического, σ – стандартное отклонение, U – значение критерия Манна-Уитни, p – вероятность ошибки. Значимые различия U критерия – полужирный шрифт.
Anthericum ramosum характеризуется низким покрытием в травостое полидоминантных остепненных лугов с одиночными генеративными деревьями. Под пологом деревьев плотность ценопопуляции венечника резко снижается – всего 7 особей на 1 м2. Это в восемь раз ниже, чем в полидоминантных остепненных лугах. Здесь формируется незавершенный левосторонний онтогенетический спектр с максимумом на v и gн особях (рис. 6, 2б). В ценопопуляции отсутствуют ss и s особи, что возможно связано с отмиранием растений уже в g3-состоянии. Кроме того, снижение плотности ценопопуляции определяется тем, что из-за небольшого светового довольствия возрастает смертность j и im особей. Группа v и gн особей пополняется за счет единичных временно нецветущих особей и партикул, формирующихся в результате дезинтеграции g2 растений. По этой же причине незначительно возрастает число g3 особей, которые представлены ветвящимися и неветвящимися партикулами.

Рисунок 10. Результаты DCA–ординации геоботанических описаний сообществ степных растений в осях наибольшего варьирования флористического состава. Сообщества: 1 – полидоминантные остепненные луга, 2 – полидоминантные остепненные луга с одиночными генеративными деревьями
Проективное покрытие Anemone sylvestris значительно меньше, чем в полидоминантных остепненных лугах. Плотность ценопопуляции уменьшается в шесть раз – всего 12 особей на 1 м2. В тени большинство особей ветреницы не формируют цветоносы. Поэтому генеративная фракция представлена единичными плодоносящими особями, среди которых преобладают g1 растения. Они в основном встречаются на периферии крон, где есть боковая подсветка. В результате плотность генеративных растений меньше в десять раз, чем в предыдущем сообществе. Это приводит к значительному сокращению пополнения ценопопуляции молодыми растениями, поскольку снижается численность g2 особей, которые дают наибольшее число корневых отпрысков. Несмотря на это онтогенетический спектр ветреницы остается полночленным левосторонним. Однако он отличается чрезвычайно низким участием генеративных особей и смещением максимума на v растения (рис. 6, 3б). Последнее связано с относительно большой длительностью v-онтогенетического состояния и незначительным пополнением ценопопуляции j и im особями.
Заключение
Максимальное видовое разнообразие полидоминантных остепненных лугов поддерживается на крутых склонах, где невозможна распашка, затруднены сенокошение и выпас, а также нечасто случаются пожары. Это способствует формированию устойчивых ценопопуляций модельных видов. Их онтогенетические спектры относятся к одному типу – полночленному левостороннему, максимум в котором приходится на молодые особи. Механизм образования этого спектра видоспецифичен. Так, особям Iris aphylla и Anthericum ramosum свойственно вегетативное размножение с неглубоким омоложением партикул и частые перерывы в цветении. Кроме того, левосторонняя структура венечника обеспечивается высокой продуктивностью семян, а Anemone sylvestris – активным вегетативным размножением, при котором особи глубоко омолаживаются. Одиночные деревья (Quercus robur, Tilia cordata) неоднозначно влияют растительность полидоминантных остепненных лугов. С одной стороны, с появлением деревьев возрастает видовое разнообразие сообществ. Это связано с тем, что деревья – удобные места отдыха и укрытия для птиц, которые разносят диаспоры растений. С другой стороны, взрослые деревья затеняют травяной покров. Это приводит к сокращению покрытия и встречаемости степных и сухолуговых видов, а также влияет на их популяционную структуру. Численность особей всех онтогенетических состояний значительно снижается. Онтогенетический спектр Anemone sylvestris остается полночленным, Iris aphylla становится неполночленным, а Anthericum ramosum – незавершенным. Если со временем число деревьев на склоне увеличится, и они сформируют сомкнутый участок леса, то ценопопуляции модельных видов постепенно исчезнут из сообщества.
Благодарности
Работа выполнена в рамках темы госзадания ЦЭПЛ РАН «Методические подходы к оценке структурной организации и функционирования лесных экосистем» (номер государственной регистрации АААА-А18-118052400130-7). Авторы выражают благодарность с.н.с. ЦЭПЛ РАН Гаврилюку Е.А. за помощь в оформлении карты.
Список литературы
Cramp S. The complete birds of the Western Palearctic. UK, 1998. CD-ROM.
Dauber J., Wolters V. Microbial activity and functional diversity in the mounds of three different ant species. // Soil Biol. Biochem. 2000. Vol. 32. Issue 1. P. 93-99.
Kostrakiewicz K. Wplyw zwierzat i drobnoustrojуw na populacje kosaccуw // Chronmy Przyr. Ojczysta. 2004. Vol. 60. No 2. P. 34-42.
Manning A., Fischer J., Lindenmayer D.B. Scattered trees are keystone structures – Implications for conservation // Biol. cons. 2006. No 132. P. 311-321.
Prevedello J.A., Almeida-Gomes M., Lindenmayer D.B. The importance of scattered trees for biodiversity conservation: A global meta-analysis // Journal of Applied Ecology. 2018. No 55. P. 205-214.
The Plant List. URL: http://www.theplantlist.org/ (дата обращения 15.10.2010).
Аверинова Е.А. Остепнённые опушечные сообщества памятников природы «Меловицкие Склоны» и «Урочище Печное» (Комаричский район Брянской области) // Изучение и охрана биологического разнообразия Брянской области: материалы по ведению Красной книги Брянской области. Вып. 5. Брянск, 2010. С. 21-26.
Барыкина Р.П., Потапова Н.Ф. Биоморфологический анализ видов рода Anemone L. флоры бывшего СССР в ходе онтогенеза // Бюл. МОИП. Отд. биол. 1994. Т. 99. Вып. 5. С. 124-137.
Барыкина Р.П., Потапова Н.Ф. Биоморфологический анализ видов рода Anemone L. флоры бывшего СССР в ходе онтогенеза // Бюл. МОИП. Отд. биол. 1994. Т. 99. Вып. 5. С. 124-137.
Босек П.З. О распространении степных растений на территории Брянской области // Бот. журн. 1980. Т. 65. № 6. С. 829–836.
Булохов А.Д. Степные элементы во флоре Брянской области // Бот. журн. 1977. № 10. С. 1505-1511.
Булохов А.Д. Травяная растительность Юго-Западного Нечерноземья России. Брянск, Изд-во БГПУ, 2001. 296 с.
Булохов А.Д., Величкин Э.М. Определитель растений Юго-Западного Нечерноземья России (Брянская, Калужская, Смоленская, Орловская области). Брянск: Изд-во БГПУ, 1997. 320 с.
Восточноевропейские широколиственные леса / Под ред. О.В. Смирновой. М.: Наука, 1994. 364 с.
Горнов А.В., Панасенко Н.Н., Комарова М.В., Тарасенко А.В. Некоторые особенности популяционной биологии Anemone sylvestris L. (Ranunculaceae) в Брянской области // Бюллетень Брянского отделения РБО. 2013. № 1(1). С. 25-30.
Горнова М.В., Евстигнеев О.И. Онтогенез и состояние ценопопуляций Melandrium dioicum (Cariophyllaceae) в высокотравных ельниках зоны широколиственных лесов (Брянская область) // Бот. журн. 2016. Т. 101. № 8. С. 896-910.
Джонгман, Р.Г.Г., Тер Браак С.Дж.Ф., Ван Тонгерен О.Ф.Р. Анализ данных в экологии сообществ и ландшафтов. М.: РАСХН, 1999. 306 с.
Евстигнеев О.И., Ручинская Е.В., Горнов А.В. Изменение остепненных лугов в широколиственно-лесной зоне под воздействием палов и хозяйственной деятельности (Брянская обл.) // Бот. журн. 2018а. Т. 103. № 12. С. 1552-1564.
Евстигнеев О.И., Ручинская Е.В., Горнов А.В. Онтогенез и состояние ценопопуляций Iris aphylla (Iridaceae) в Брянской области // Бот. журн. 2018б. Т. 103. № 2. С. 207-223.
Евстигнеев О.И., Федотов Ю.П., Горнов А.В. К флоре памятника природы «Севские склоны» // Изучение и охрана биологического разнообразия Брянской области. Материалы по ведению Красной книги Брянской области. Брянск, 2011. Вып. 6. С. 45-52.
Евстигнеев О.И., Федотов Ю.П., Кайгородова Е.Ю. Природа Неруссо-Деснянского полесья Брянской области. Редкие растения. Брянск, 2000. 223 с.
Евстигнеев О.И., Харлампиева М.В. Онтогенез и состояние популяций Ligularia sibirica (Asteraceae) в ненарушенных ельниках на низинных болотах (Брянская область) // Бот. журн. 2014. Т. 99. № 6. С. 670-681.
Жукова Л.А. Изменение возрастного состава популяций луговика дернистого на Окских лугах: Автореф. дисс. … канд. биол. наук (спец. 03.02.01). М.: МПГИ им. В.И. Ленина, 1967. 19 с.
Журавлева Е.Н., Ипатов В.С., Лебедева В.Х., Тиходеева М.Ю. Изменение растительности на лугах под влиянием сосны обыкновенной (Pinus sylvestris L.) // Вестник Санкт–Петербургского университета. 2012. Сер. 3. Вып. 2. С. 3-12.
Заугольнова Л.Б. Методика сбора и объем материала // Восточноевропейские широколиственные леса. М.: Наука. 1994. C. 74-93.
Заугольнова Л.Б. Структура популяций семенных растений и проблемы их мониторинга: Дис. … докт. биол. наук (спец.03.00.05). СПб: СПБГУ, 1994. 70 с.
Заугольнова Л.Б., Смирнова О.В. Возрастная структура ценопопуляций многолетних растений и ее динамика // Журн. общ. биол. 1978. Т. 39. № 6. С. 849-858.
Заугольнова Л.Б., Смирнова О.В., Комаров А.С., Ханина Л.Г. Мониторинг фитопопуляций // Успехи соврем. биол. 1993. Т. 113. № 4. С.402-414.
Зеленая книга Брянской области (растительные сообщества, нуждающиеся в охране). Брянск: «Брянский государственный университет имени академика ИГ Петровского», 2012. 144 с.
Зрянин В.А. Влияние муравьев рода Lasius на почвы луговых биогеоценозов // Успехи совр. биологии. 2003. Т. 123. № 3. С. 278-287.
Ипатов В.С. Фитогенные поля одиночных деревьев некоторых пород в одном экотопе // Бот. журн. 2007. Т. 92. № 8. С. 1186-1192.
Комаров Н.Ф. Этапы и факторы эволюции растительного покрова черноземных степей. М., 1951. 328 с.
Красная книга Брянской области. 2-е издание. Брянск: РИО БГУ, 2016. 432 с.
Красная книга Брянской области. Растения, грибы. Брянск: ЗАО Изд-во «Читай-город», 2004. 272 с.
Красная книга Калужской области. Калуга, ООО «Ваш домъ», 2015. Т. 1. 536 с.
Красная книга Курской области. Редкие и исчезающие виды растений и грибов. Тула, 2002. Т. 2. 165 с.
Красная книга Российской Федерации (растения и грибы). М.: Товарищество научных изданий КМК, 2008. 855 с.
Левина Р.Е. Способы распространения плодов и семян. М.: Изд-во МГУ. 1957. 361 с.
Миркин Б.М., Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов современной фитоценологии. М.: «Наука», 1989. 223 с.
Никонов В.В., Лукина Н.В., Смирнова Е.В., Исаева Л.Г. Влияние Picea obovata и Pinus sylvestris на первичную продуктивность нижних ярусов хвойных лесов Кольского полуострова // Бот. журн. 2002. Т. 87. № 8. С. 107-119.
Одум Ю. Экология. Т. 2. М.: «Мир», 1986. 376 с.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Артемкина Н.А. Влияние ели на кислотность и содержание элементов питания в почвах северотаежных ельников кустарничково-зеленомошных // Почвоведение. 2016. № 11. С. 1355-1367.
Панасенко Н.Н., Евстигнеев О.И., Горнов А.В., Ручинская Е.В. К флоре памятника природы «Меловицкие склоны» (Брянская область) // Бюллетень Брянского отделения РБО. 2015. Т. 2. № 6. С. 17-25.
Панасенко Н.Н., Евстигнеев О.И., Федотов Ю.П., Горнов А.В. К флоре памятника природы «Марковские горы» (Брянская область) // Изучение и охрана биологического разнообразия Брянской области. Материалы по ведению Красной книги Брянской области. Вып. 8. Брянск, 2013. С. 121-131.
Природное районирование и типы сельскохозяйственных земель Брянской области. Брянск, Приок. кн. изд-во. Брян. отд., 1975. 611 с.
Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Труды БИН АН СССР. Серия 3. Геоботаника. М.-Л., 1950. № 6. С. 7-204.
Растительность Европейской части СССР. 1980. Л.: «Наука», 1980. 420 с.
Ручинская Е.В. Структурное и видовое разнообразие растительности остепненных лугов в зоне широколиственных лесов (на примере памятника природы «Меловицкие склоны», Брянская обл.): Дисс. … канд. биол. наук (спец. 03.02.08). М., 2019. 197 с.
Самойлов Ю.И. Структура фитогенного поля на примере одиночных дубов Quercus robur (Fagaceae) // Бот. журн. 1983. Т. 68. № 8. С. 1022-1034.
Семенищенков Ю.А. Кальцефитная травяная растительность в Брянской области: синтаксономия, экология и вопросы охраны // Вестник ВГУ. Серия: Химия. Биология. Фармация. 2012. № 1. С. 149-163.
Семенищенков Ю.А. Остепненные луга правобережья реки Десны – уникальные природные комплексы на границе ботанико-географических подзон хвойно-широколиственных и широколиственных лесов // Проблемы изучения и восстановления ландшафтов лесостепной зоны. Тула, 2010. Вып. 1. С. 206-216.
Серебряков И.Г. Экологическая морфология растений. М.: «Высшая школа», 1962. 378 с.
Скворцов А.К. Кальцефильная флора на юге Погарского района Брянской области // Бюлл. МОИП. Отд. биол. 1982. Т. 87. Вып. 5. С. 77-83.
Старостенкова М.М. Род ветреница / В кн.: Биол. флора Московской области. М.: Издательство Московского университета, 1976. Вып. 3. С. 119-138.
Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов // Научные доклады высшей школы. Биологические науки. 1975. № 2. С. 7-34.
Уранов А.А. Фитогенное поле // Проблемы современной ботаники. Т. 1. М.-Л: «Наука», 1965. С. 251-254.
Ценопопуляции растений (очерки популяционной биологии). М.: «Наука», 1988. 184 с.
Чернова Н.М., Былова А.М. Общая экология. М.: Дрофа, 2004. 416 с.
Рецензент: к.б.н., доцент Панасенко Н.Н.
Приложение
Видовой состав сообществ памятника природы «Меловицкие склоны»
| Название растения | БВ | ЭЦГ | |
| Сообщества | |||
| 1 | 2 | ||
| Achillea millefolium L. | IV (+) | II (+) | Су-Лу |
| Agrimonia eupatoria L. | V (+) | IV(+) | Су-Лу |
| Ajuga genevensis L. | I (+) | I (+) | Су-Лу |
| Allium oleraceum L. | – | I (+) | Су-Лу |
| Anemone sylvestris L. | V (1) | II (1) | Су-Лу |
| Anthericum ramosum L. | V (1) | V (+) | Су-Лу |
| Anthyllis vulneraria L. | I (+) | – | Су-Лу |
| Artemisia absinthium L. | – | I (+) | Су-Лу |
| Artemisia vulgaris L. | – | II (+) | Су-Лу |
| Asparagus officinalis L. | I (+) | V (+) | Су-Лу |
| Aster amellus L. | V (3) | II (1) | Су-Лу |
| Astragalus cicer L. | IV (+) | V (1) | Су-Лу |
| Astragalus glycyphyllos L. | I (+) | IV (+) | Су-Лу |
| Brachypodium pinnatum (L.) Beauv. | II (2) | V (3) | Не-Оп |
| Bromus inermis Leyss. | V (3) | V (4) | Су-Лу |
| Calamagrostis epigejos (L.) Roth | IV (+) | V (1) | Су-Лу |
| Campanula bononiensis L. | V (+) | V (+) | Су-Лу |
| Campanula rapunculoides L. | II (+) | II (+) | Су-Лу |
| Campanula sibirica L. | II (+) | – | Су-Лу |
| Carex hirta L. | – | I (+) | Вл-Лу |
| Carex lachenalii Schkuhr | – | IV (+) | Вл-Лу |
| Carex montana L. | – | II (+) | Су-Лу |
| Carex praecox Schreb. | III (+) | IV (+) | Су-Лу |
| Centaurea jacea L. | III (+) | – | Су-Лу |
| Centaurea phrygia subsp. pseudophrygia (C.A. Mey.) Gugler | III (1) | II (+) | Су-Лу |
| Chamaecytisus ruthenicus (Fisch. ex Woloszcz.) Klaskova | V (+) | III (+) | Су-Лу |
| Cichorium intybus L. | I (+) | – | Су-Лу |
| Cirsium decussatum Janka | – | I (+) | Су-Лу |
| Cirsium pannonicum (L. fil.) Link | IV (1) | III (+) | Су-Лу |
| Convallaria majalis L. | IV (1) | IV (1) | Не-Ле |
| Convolvulus arvensis L. | III (+) | V (+) | Су-Лу |
| Corylus avellana L. | IV (+) | V (+) | Не-Ле |
| Dactylis glomerata L. | III (+) | V (+) | Вл-Лу |
| Elymus repens (L.) Gould | II (+) | II (+) | Су-Лу |
| Equisetum arvense L. | – | I (+) | Су-Лу |
| Erigeron annuus (L.) Desf. | – | I (+) | Су-Лу |
| Euonymus europaeus L. | – | III (+) | Не-Ле |
| Euphorbia esula L. | II (+) | I (+) | Су-Лу |
| Euphorbia semivillosa (Prokh.) Krylov | V (2) | V (+) | Су-Лу |
| Fallopia convolvulus (L.) Á. Löve | – | I (+) | Су-Лу |
| Festuca pratensis Huds. | II (+) | – | Су-Лу |
| Filipendula vulgaris Moench | – | IV (+) | Су-Лу |
| Fragaria viridis Weston | III (+) | III (+) | Су-Лу |
| Frangula alnus Mill. | IV (+) | II (+) | Бо-Ле |
| Galatella linosyris (L.) Rchb.f. | II (+) | II (+) | Су-Лу |
| Galeopsis bifida Boenn. | – | III (+) | Вл-Лу |
| Galium aparine L. | – | IV (+) | Че-Оп |
| Galium boreale L. | V (+) | V (1) | Су-Лу |
| Galium mollugo L. | V (1) | V (1) | Вл-Лу |
| Galium tinctorium L. | V (1) | V (+) | Су-Лу |
| Galium verum L. | V (+) | V (+) | Су-Лу |
| Genista tinctoria L. | IV (+) | I (+) | Су-Лу |
| Geranium sanguineum L. | IV (+) | IV (+) | Су-Лу |
| Hypericum perforatum L. | I (+) | III (+) | Су-Лу |
| Inula hirta L. | II (1) | IV (+) | Су-Лу |
| Inula salicina L. | V (1) | V (+) | Су-Лу |
| Iris aphylla L. | V (3) | V (2) | Су-Лу |
| Knautia arvensis (L.) Coult. | III (+) | V (+) | Су-Лу |
| Lactuca serriola L. | – | I (+) | Су-Лу |
| Laserpitium latifolium L. | IV (1) | V (2) | Не-Оп |
| Lathyrus niger (L.) Bernh. | – | V (1) | Не-Ле |
| Lathyrus pisiformis L. | I (+) | – | Не-Оп |
| Lathyrus sylvestris L. | II (+) | IV (1) | Не-Оп |
| Lavatera thuringiaca L. | I (+) | I (+) | Су-Лу |
| Leucanthemum vulgare (Vaill.) Lam. | IV (1) | I (+) | Су-Лу |
| Linum flavum L. | IV (+) | I (+) | Су-Лу |
| Lithospermum officinale L. | IV (+) | IV (+) | Су-Лу |
| Medicago falcata L. | I (+) | V (+) | Су-Лу |
| Myosotis ramosissima Rochel | – | I (+) | Су-Лу |
| Nepeta nuda L. | I (1) | IV (2) | Су-Лу |
| Origanum vulgare L. | IV (1) | V (1) | Су-Лу |
| Peucedanum alsaticum L. | V (1) | V (+) | Су-Лу |
| Peucedanum cervaria (L.) Cusson ex Lapeyr. | IV (1) | V (1) | Не-Оп |
| Peucedanum oreoselinum (L.) Moench | I (+) | – | Су-Лу |
| Phlomoides tuberosa (L.) Moench | II (+) | II (1) | Су-Лу |
| Pilosella piloselloides subsp. bauhinii (Schult.) S.Bräut. & Greuter | I (+) | – | Су-Лу |
| Plantago lanceolata L. | I (+) | – | Су-Лу |
| Plantago media L. | I (+) | – | Су-Лу |
| Poa angustifolia L. | V (1) | V (1) | Су-Лу |
| Poa trivialis L. | – | IV (+) | Вл-Лу |
| Podospermum purpureum (L.) W.D.J. Koch & Ziz | I (+) | – | Су-Лу |
| Polygala comosa Schkuhr | III (+) | – | Су-Лу |
| Polygonatum odoratum (Mill.) Druce | III (+) | II (+) | Су-Лу |
| Prunus cerasus L. | IV (1) | III (+) | Су-Лу |
| Pteridium aquilinum (L.) Kuhn | IV (2) | V (2) | Бо-Оп |
| Pyrethrum corymbosum (L.) Scop. | V (1) | V (1) | Не-Оп |
| Pyrus communis L. | – | I (+) | Не-Ле |
| Quercus robur L. | I (+) | V (+) | Не-Ле |
| Ranunculus polyanthemos L. | IV (+) | I (+) | Су-Лу |
| Rubus caesius L. | II (+) | III (1) | Че-Оп |
| Salvia pratensis L. | V (3) | V (1) | Су-Лу |
| Salvia verticillata L. | II (+) | I (+) | Су-Лу |
| Securigera varia (L.) Lassen | IV (1) | V (1) | Су-Лу |
| Sedum maximum (L.) Suter | I (+) | – | Су-Лу |
| Sedum telephium L. | I (+) | – | Су-Лу |
| Serratula tinctoria L. | – | I (+) | Не-Оп |
| Seseli libanotis (L.) W.D.J. Koch | I (2) | III (+) | Су-Лу |
| Silene latifolia Poir. | I (+) | III (+) | Су-Лу |
| Silene nutans L. | I (+) | I (+) | Су-Лу |
| Silene viscaria (L.) Jess. | – | I (+) | Су-Лу |
| Silene vulgaris (Moench) Garcke | – | I (+) | Су-Лу |
| Solidago virgaurea L. | I (+) | III (+) | Бо-Оп |
| Stachys officinalis (L.) Trevis. | – | V (+) | Су-Лу |
| Stachys recta L. | V (3) | V (1) | Су-Лу |
| Succisa pratensis Moench | I (+) | – | Вл-Лу |
| Taraxacum officinale Wigg. | II (+) | I (+) | Су-Лу |
| Thalictrum lucidum L. | III (+) | – | Вл-Лу |
| Thalictrum minus L. | III (1) | III (+) | Су-Лу |
| Tilia cordata Mill. | – | IV (+) | Не-Ле |
| Tragopogon dubius Scop. | I (+) | – | Су-Лу |
| Trifolium alpestre L. | V (1) | V (1) | Су-Лу |
| Trifolium montanum L. | V (+) | II (+) | Су-Лу |
| Valeriana officinalis L. | III (+) | II (+) | Че-Оп |
| Verbascum lychnitis L. | II (+) | I (+) | Су-Лу |
| Verbascum nigrum L. | V (+) | IV (+) | Су-Лу |
| Veronica austriaca subsp. teucrium (L.) D.A. Webb | V (1) | V (1) | Су-Лу |
| Veronica chamaedrys L. | – | I (+) | Су-Лу |
| Veronica spuria L. | – | I (+) | Су-Лу |
| Vicia tenuifolia Roth | V (2) | V (1) | Су-Лу |
| Vicia tetrasperma (L.) Schreb. | – | I (+) | Вл-Лу |
| Vincetoxicum hirundinaria Medik. | V (1) | V (+) | Су-Лу |
| Viola canina L. | I (+) | – | Вл-Лу |
| Viola collina Besser. | III (+) | IV (+) | Бо-Оп |
| Viola hirta L. | IV (+) | V (+) | Су-Лу |
| Viola mirabilis L. | I (+) | III (+) | Не-Ле |
| Число видов | 98 | 107 | |
Примечание. Сообщества: 1 – полидоминантные остепненные луга, 2 – полидоминантные остепненные луга с одиночными генеративными деревьями. БВ – средние баллы встречаемости, «+» и арабские цифры – баллы покрытия-обилия по шкале Браун-Бланке. ЭЦГ – эколого-ценотические группы: Су-Лу – сухолуговая, Вл-Лу – влажно-луговая, Бо-Оп – боровая (бореальная опушечная), Не-Оп – неморальная опушечная, Не-Ле – неморальная лесная, Че-Оп – черноольховая опушечная, Бо-Ле – бореальная лесная.





