- DOI 10.31509/2658-607x-202252-109
- УДК 630*884+630*114.6+630*181.9
Сравнительная оценка сукцессионной динамики древесных фракций ельников южной тайги
![]()
© 2022 г. В. Г. Стороженко
Институт лесоведения РАН
Россия, 143030, Московская область, с. Успенское, ул. Советская, д. 21
E-mail: lesoved@mail.ru
Поступила в редакцию: 05.04.2022
После рецензирования: 01.08.2022
Принята к печати: 10.08.2022
Актуальность. Коренные леса можно признать исчезающими формациями Европейского Севера России. Они являются эталонами устойчивости лесов для сравнительной оценки с лесами другого происхождения и использования, что определяет актуальность исследований их структурных особенностей. Цель исследований — провести сравнительную оценку динамических процессов в коренных разновозрастных лесах еловых формаций подзоны южной тайги различных сукцессионных положений (фаз динамики) во временном тренде их развития.
Материал и методы. Исследованы коренные разновозрастные ельники подзоны южной тайги Центрально-Лесного биосферного государственного заповедника (ЦЛБГЗ) (Тверская обл.) и заповедника «Кологривский лес» (Костромская обл.) различных динамических характеристик и сукцессионного положения — климаксных, демутационных, дигрессивных. В биогеоценозах пробных площадей определялся возраст деревьев, присутствие гнилей, строились возрастные ряды, вычислялись объемы деревьев в возрастных поколениях. В один временной ряд выстраивались все древесные фракции анализируемых ельников: древостоев, текущего древесного отпада и валежа — от ретроспективных значений (– 60 лет) и до перспективных значений предельного возраста деревьев первых поколений древостоев (+ ~ 300 лет). Описаны процессы динамики объемов древесной фракции коренных ельников различных сукцессионных положений в длительном временном пространстве с различной скоростью протекания процессов накопления и разложения древесной биомассы.
Результаты и обсуждение. Процессы накопления биомассы и формирования возрастных структур биогеоценозов идут в 4–7 раз медленнее, чем процессы разложения отмирающей биомассы древесного отпада. Дереворазрушающие грибы сапротрофного комплекса «подстраивают» активность процесса разложения древесного отпада под динамику накопления древесины древостоем, сохраняя баланс накопления и разложения древесной биомассы. Корреляционная зависимость увеличения значений пораженности деревьев дереворазрушающими грибами в возрастных поколениях с увеличением их возраста выражается в коэффициентах корреляции от r — 0.89 при mr — 0.07 и t — 11.8 (ельник 1) до 0.99 при mr — 0.004 и t — 245 (ельник 4). Связь в обоих случаях очень высокая, почти функциональная, которая может трактоваться как закономерность.
Заключение. В коренных девственных лесах сохраняется баланс воспроизводимой и разлагаемой древесины, и этот баланс служит одним из важнейших критериев устойчивого функционирования лесных сообществ. Это положение можно расценивать как важный факт коэволюционной функциональной структуры лесов.
Ключевые слова: южная тайга, коренные еловые леса, возрастные поколения, древесные фракции, баланс биомассы
Изучение динамических процессов в коренных лесных биогеоценозах можно признать одним из краеугольных положений лесной науки, на основе которых строятся фундаментальные теории сукцессионных законов развития лесных сообществ, переходящих в практику формирования лесов различного назначения и использования (Морозов, 1962; Сукачев, 1964; Исаев и др., 2008; Tikkanen at al., 2014; Стороженко, 2007, 2011). Коренные леса эволюционного формирования, не затронутые антропогенным воздействием, в нашей современной реальности можно признать исчезающими формациями на всем пространстве Европейского Севера России. Именно леса такого статуса являются эталонами устойчивости как качества структурного строения сообществ, выработанного в постоянной борьбе за восстановление оптимальных связей между консортами ценозов, нарушенными различными абиотическими факторами — пожарами, ветровалами, климатическими аномалиями и т. д. Восстановительная динамика лесов на разных этапах сукцессий оригинальна в каждом отдельном случае и определяется огромным количеством условий развития как автотрофных, так и гетеротрофных сообществ, входящих в структуру и функциональную организацию биогеоценозов на уровне видов, сообществ разного таксономического и ценотического уровней.
С этих позиций представляется актуальным изучение процессов трансформации древесной биомассы коренных девственных биогеоценозов на разных временных этапах динамики в сукцессионном тренде их развития — от ретроспективы в 40–50 лет через настоящий период времени и перспективу до возраста первых поколений возрастных рядов.
Из всех лесных формационных возможностей для поставленных целей более всего подходят леса еловых формаций, менее других подверженные пирогенным воздействиям, формирующие сложные возрастные и горизонтальные и возобновительные структуры.
Цель исследований — провести сравнительную оценку динамических процессов в коренных разновозрастных лесах еловых формаций подзоны южной тайги различных сукцессионных положений (фаз динамики) во временном тренде их развития.
МАТЕРИАЛ И МЕТОДЫ
В качестве объектов исследований приняты коренные разновозрастные ельники Центрально-Лесного биосферного государственного заповедника (ЦЛБГЗ) (Тверская обл.) (56°26´…56°31´ с. ш.; 32°29´…33°29´ в. д.) и заповедника «Кологривский лес» (Костромская обл.), (58°56′41″ с. ш.; 43°51′03″ в. д.), различных динамических характеристик, сукцессионного положения: климаксных, демутационных, дигрессивных, входящих в подзону южной тайги (Курнаев, 1973). Постоянные пробные площади (ППП) закладывались в ельниках наиболее производительной кисличной группы типов леса. Размеры ППП составляли от 0.4 до 0.6 га. На пробных площадях проводилось лесоводственное описание биогеоценозов, деревья нумеровались и картировались в горизонтальной проекции. Стволы деревьев бурились у шейки корня возрастным буром Пресслера с определением их возраста и наличия гнилевых фаутов с фиксацией диаметра и типа гнилей (коррозионные и деструктивные) (Стороженко и др., 1992, 2011). Проводился учет естественного возобновления всех пород в градациях высоты через 0.5 м и определением возраста по мутовкам. На площади участков учитывались валежные стволы всех пород с измерением диаметров, длины стволов, стадии разложения (Стороженко, 1990). Все деревья относились к определенной категории состояния по принятой в лесопатологии шестибальной шкале (Правила…, 2013). На площади участков и в прилегающих ельниках близких условий роста собирали плодовые тела грибов дереворазрушающего комплекса как биотрофов, так и сапротрофов с последующей идентификацией грибов по определителям (Бондарцева, 1998; Ryvarden, Gilbertson, 1993, 1994; Niemelä, 2001, 2005). В определении грибов неоценимую помощь оказала В. М. Коткова (БИН РАН).
В камеральный период определялись объемные показатели деревьев, запасы древостоев, объемы валежных структур и текущего древесного отпада (Третьяков и др., 1952), строились возрастные ряды древостоев биогеоценозов по сорокалетним возрастным поколениям, ряды валежных структур по стадиям разложения и текущего древесного отпада по категориям состояния. Сукцессионное положение биогеоценозов (фаза динамики) определяется по суммарному соотношению числа деревьев или, как в нашем случае, объемов деревьев в возрастных поколениях от среднего возрастного поколения возрастного ряда древостоя (Дыренков, 1984; Стороженко, 2007).
Таким образом, в один временной ряд выстраивались все древесные фракции анализируемых лесных сообществ — от ретроспективных значений (–60 лет) и до перспективных значений предельного возраста деревьев первых поколений древостоев (+ ~300 лет). Для анализа связи присутствия гнилей в деревьях с увеличением их возраста применен коэффициент корреляции по качественному признаку.
Полученные сведения позволили в экспериментальном формате оценить процессы движения объемов древесной фракции коренных ельников различных сукцессионных положений в длительном временном пространстве с различной скоростью протекания процессов накопления и разложения древесной биомассы.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Коренные ельники Кологрива расположены примерно на одной широте с ельниками ЦЛБГЗ, но в шестистах километрах восточнее в их составе присутствует пихта, занимающая на этой широте в естественных лесах самое западное расположение. В табл. 1 приведены основные лесоводственные характеристики изучаемых биогеоценозов.
Таблица 1. Лесоводственные характеристики коренных разновозрастных ельников южной тайги
| № ППП | Лесоводственные характеристики: состав, тип леса, полнота, бонитет | Запас,
м3/га |
Средний диаметр,
см |
Средняя высота, м | Средний возраст, лет | Подрост | Подле-
сок |
Покров |
| Ельники Центрально-Лесного биосферного заповедника | ||||||||
| 1 | 9Е1Ос + Б, кис-чер, 0.7, I |
581.6 | 33.9 | 30.5 | 163.4 | Е, Б, Ос | Рб, Мж | Кис, чер, мхи, пап |
| 2 | 8Е2Ос + Б, чер-пап, 0.8, I | 408.8 | 26.1 | 27.0 | 140.2 | Е, Ос, Б | Рб, Мж | Кис, чер, мхи, пап |
| Ельники заповедника «Кологривский лес» | ||||||||
| 3 | 8Е1Б1Лп, кис-щит, 0.8, I | 396.5 | 25.9 | 26.5 | 155.3 | Е, Пх, Лп | Рб, Мж, | Кис, чер, мхи, пап |
| 4 | 10Е + Пх,Б; кис-пап, 0.8, I | 438.8 | 20.7 | 24.5 | 144.2 | Е, Пх, Б | Рб,Мж | Кис, чер, мхи, пап |
Обозначения. Типы леса: кис-чер — кислично-черничный, чер-пап — чернично-папоротниковый, кис-щит — кислично-щитовниковый, кис-пап — кислично-папоротниковый.
Из данных таблицы 1 видно: несмотря на то, что бонитеты всех четырех ельников одинаково высоки, их запасы, средние высоты и средние диаметры различаются, что объясняется различными средними возрастами, то есть числом деревьев в разных возрастных поколениях в древостоях. Можно отметить также присутствие в лесах «Кологривского леса» некоторой примеси пихты в составе древостоя и подроста как представителя более восточных и северных лесов Европейской России, а также общие меньшие линейные параметры деревьев, что, однако, ненамного снижает высокие показатели их производительности.
Более детальный анализ структурных особенностей изучаемых ельников приведен в таблице 2.
Древостой биогеоценоза 1 сложен семью возрастными поколениями, и среднее положение в возрастном ряду занимает четвертое поколение. Для вычисления объемов деревьев в восходящих и нисходящих рядах объемы среднего поколения делятся пополам. Таким образом, по объемным показателям в поколения восходящего ряда входят 201.3 м3/га объема древесины, нисходящего ряда — 380.3 м3/га. В четвертом поколении сосредоточен наибольший объем древесины, и по методике И. И. Гусева (1964) и С. А. Дыренкова (1984) он близок к фазе климакса. В то же время большие объемы древесины, сосредоточенные в поколениях нисходящего ряда, относят его к сообществам, прошедшим эту фазу и двигающимся к фазе дигрессии. Таким образом, биогеоценоз в динамике развития более отвечает статусу климаксово-дигрессивного. Динамика объемов первых двух поколений возрастного ряда и старого сухостоя объясняет большие объемы валежа первых двух стадий разложения, перешедших в валеж в последние 10–20 лет. Очень высокие значения объемов старого сухостоя в составе текущего древесного отпада подтверждают этот вывод.
Древостой биогеоценоза 2, так же как и древостой 1, имеет семь возрастных поколений и находится очень близко к состоянию климакс на пути из демутации, освобождаясь в течение последних 30–40 лет от значительных объемов древесины, переходящей в структуру валежа (2–4 стадии разложения). При этом значения объемов ТДО малы, что связано с некоторой стабилизацией динамического процесса при вхождении в фазу климакса. Биогеоценоз, следовательно, в динамике развития можно трактовать как демутационно-климаксовый.
Биогеоценоз 3 имеет восемь возрастных поколений и, таким образом, середина возрастного ряда приходится на границу четвертого и пятого поколений. В таком случае для расчета объемов середины возрастного ряда объем пятого поколения распределяется поровну между четвертым и шестым возрастным поколением. Объемы восходящего ряда составляют 85.85 м3/га, а нисходящего — 310.65 м3/га. Понятно, что биогеоценоз склоняется к фазе дигрессии, и это движение сохранится долгое время, пока объемы четвертого и пятого поколений будут двигаться по сукцессионной лестнице к предельным возрастам сообщества. В то же время структура возрастного ряда отличается довольно равномерным распределением объемов деревьев в возрастных поколениях, что характерно для сообществ, находящихся в относительно длительной динамической стабилизации, то есть близкой к климаксу. Об этом же свидетельствует и относительно равномерное поступление деревьев в структуру валежа по стадиям разложения. С некоторой осторожностью биогеоценоз можно отнести к климаксово-дигрессивной фазе динамики.
Близок к биогеоценозу 3 биогеоценоз 4, который, однако, имеет большие объемы древесины нисходящего ряда, из возрастных поколений которого постоянно пополняются объемы валежа, особенно в ретроспективе до 50 лет (пятая стадия разложения). Биогеоценоз 4 более близок к дигрессивной фазе динамики, чем биогеоценоз 3.
Таблица 2. Параметры древесных фракций и динамические характеристики ельников южной тайги
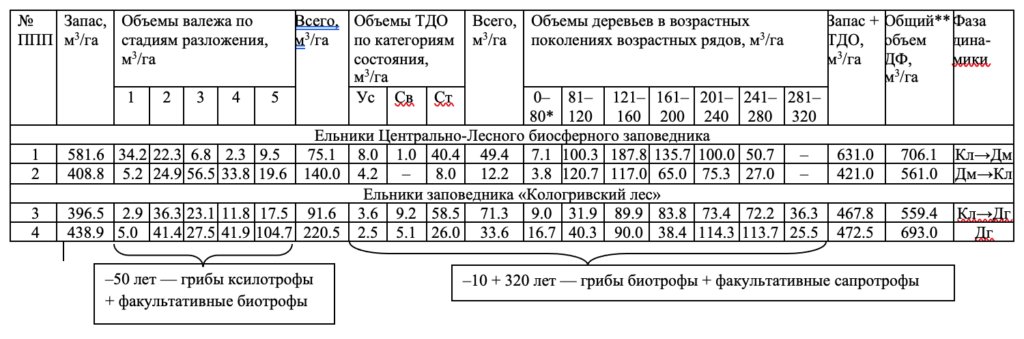
Обозначения. Фаза динамики: Кл — климакс, Дг — дигрессия, Дм — демутация. ТДО — текущий древесный отпад (категории состояния: Ус — усыхающие, Св — свежий сухостой, Ст — старый сухостой). Графа 0–80* — включает два возрастных поколения: подрост до 40 лет (последнее поколение) и 41–80 лет (предпоследнее поколение); ДФ** — древесная фракция биогеоценоза (запас древостоя + ТДО + валеж).
Динамические процессы, протекающие в представленных выше еловых сообществах, характерны для всех коренных ельников тайги и в целом для всех коренных разновозрастных лесных формаций. Древесные фракции таких лесов также сложены тремя фрагментами, отражающими этапы преобразований древесины в динамике ее трансформации от древостоя в живом состоянии до древесного отпада (табл. 2).
При этом процессы трансформации древесины одной фракции в следующую протекают в различных временных интервалах. Накопление биомассы древесины коренного разновозрастного древостоя в динамике от начала его возникновения до предельных возрастов первых поколений протекает во временном интервале от подроста до 240–320 лет для лесов еловых формаций разных подзон таежной зоны. Для этих же условий разложение древесного отпада (валежа) протекает во временном интервале от начала вывала до пятой стадии 50–60 лет. Деревья, входящие во фракцию древостоя, но относящиеся к древесному отпаду (ТДО), составляют переходную фракцию, протекающую во временном интервале до 10–15 лет. Таким образом, можно видеть большую временную разницу в протекании процессов накопления и разложения древесной биомассы от начала ее формирования до полного ее разложения. Для каждого биогеоценоза она различна и определяется делением значений предельных возрастов деревьев на период разложения древесного отпада. В нашем случае она составляет 5.6 (биогеоценозы 1 и 2) и 6.6 (биогеоценозы 3 и 4), то есть процессы накопления объемов биомассы идут медленнее, чем процессы ее разложения. В коренных девственных лесах сохраняется баланс воспроизводимой и разлагаемой древесины, и этот баланс служит одним из важнейших критериев устойчивого функционирования лесных сообществ. Это положение можно расценивать как важный факт функциональной структуры лесов.
В этой «связке» ведущей автотрофной фракцией является древостой, параметры которого определяются внешними эндогенными факторами формирования объемов биомассы — энергия Солнца, газовый состав атмосферы, водное и минеральное питание почвы. «Ведомая» фракция отдана эволюцией гетеротрофному комплексу. В нем основным разлагающим древесину агентом являются дереворазрушающие грибы, пищевым субстратом которых служат стенки клеток древесины (целлюлоза, гемицеллюлозы и лигнин) (Соловьев, 1992). Судя по представленным выше положениям, именно дереворазрушающие грибы «подстраивают» активность своего процесса разложения древесного отпада под динамику накопления древесины древостоем, сохраняя баланс накопления и разложения биомассы лесного сообщества.
Заметное участие грибов дереворазрушающего комплекса в процессах формирования структур еловых древостоев таежной зоны начинается, как правило, с возраста деревьев предпоследнего возрастного поколения и не прекращается до предельных возрастов деревьев первого поколения. В относительном выражении пораженность деревьев в пределах возрастных поколений представлена в табл. 3.
Таблица 3. Пораженность деревьев в возрастных поколениях древостоев дереворазрушающими грибами биотрофного комплекса
| № ППП | Пораженность деревьев в возрастных поколениях, % | Поражен-ность древостоя | |||||||
| До 40 | 41–80 | 81–120 | 121–160 | 161–200 | 201–240 | 241–280 | 281–320 | ||
| Ельники Центрально-Лесного биосферного заповедника | |||||||||
| 1 | Подрост | Ед | 16.2 | 24.1 | 9.0 | 9.0 | 33.0 | – | 12.9 |
| 2 | 14.8 | 20.9 | 16.7 | 21.1 | 41.2 | 48.8 | – | 18.8 | |
| Ельники заповедника «Кологривский лес» | |||||||||
| 3 | Подрост | 26.9 | 37.8 | 30.0 | 19.1 | 43.5 | 50.0 | 69.0 | 32.1 |
| 4 | 18.1 | 26.8 | 32.2 | 38.4 | 46.6 | 58.3 | 62.0 | 30.8 | |
Данные таблицы 3 демонстрируют подтвержденную ранее тенденцию возрастания значений пораженности деревьев в возрастных поколениях с увеличением их возраста (Стороженко, 2007). В изучаемых ельниках корреляционная связь этих предикторов выражается в коэффициентах корреляции от r — 0.89 при mr — 0.07 и t — 11.8 (ельник 1) до 0.99 при mr — 0.004 и t — 245 (ельник 4). Связь в обоих случаях очень высокая, почти функциональная, которая может трактоваться как закономерность.
Дереворазрушающие грибы биотрофного комплекса, поражающие живые деревья древостоя, вызывают гнили корней и стволов коррозионного и деструктивного типов, снижают их жизнеспособность, ослабляют их физические свойства и являются одной из основных причин буреломов и ветровалов. Более 80% стволов ветровала и бурелома последних 3 лет вывала имеют гнили 3 и 4 стадий, которые являются причинами их появления. Чем выше пораженность деревьев в возрастных поколениях, тем быстрее они очищаются от деревьев с развитыми гнилевыми фаутами, переходящими в древесный отпад. Именно поэтому грибы биотрофного комплекса можно признать важным эндогенным «механизмом», представляющим один из гетеротрофных консортов, регулирующих баланс биомассы коренных лесных сообществ на протяжении всего периода формирования возрастных структур древостоев.
Не менее важным гетеротрофным консортом, осуществляющим разложение древесного отпада до состояния гумуса, но со значительно более коротким временным периодом, является ксилотрофный (сапротрофный) комплекс дереворазрушающих грибов (табл. 2). Именно потому, что временные периоды участия грибов биотрофного и ксилотрофного комплексов в процессах деструкции древесины фракций древостоя и валежа различаются в несколько раз, в коренных лесах эволюционного формирования сохраняется баланс воспроизводимой и разлагаемой биомассы, что определяет их как устойчивые лесные сообщества. Важным дополнением к сказанному можно отметить присутствие в составе комплекса дереворазрушающих грибов видов, проявляющих факультативные свойства, расширяющие деструктивную функцию на весь цикл развития лесного биогеоценоза.
Видовой состав дереворазрушающих грибов очень широк (Бондарцева, 1998; Niemelä, 2005; Стороженко, Коткова, 2013; и др.), но наиболее часто встречающиеся виды биотрофного и сапротрофного комплексов, ассоциированные с лесами еловых формаций южной тайги, приведены ниже (табл. 4). Видовой состав грибов биотрофов, вызывающих гнилевые фауты живых растущих деревьев, значительно меньше, чем грибов сапротрофного комплекса, разлагающих древесный отпад.
Таблица 4. Наиболее часто встречающиеся виды дереворазрушающих грибов ельников южной тайги
| Грибы биотрофного комплекса | Тип питания | Встречае-мость | |
| Латинское название | Русское название | ||
| Armillaria borealis Marxm. et Korhonen | Опенок северный | Факультативный сапротроф | Часто |
| Climacocystis borealis (Fr.) Kotl. et Pouz. | Северный трутовик | Биотроф | Часто |
| Heterobasidion parviporum Niemelä et Korhonen | Корневая губка | Биотроф | Часто |
| Onnia triqueter (Lenz.) Imazeki | Еловая комлевая губка | Биотроф | Часто |
| Phaeolus schweinitzii (Fr.) Pat. | Трутовик Швейница | Биотроф | Часто |
| Porodaedalea chrysoloma (Fr.) Fias. et Niem. | Еловая губка | Биотроф | Часто |
| Грибы сапротрофного комплекса | |||
| Antrodia seriales (Fr.) Donk | Антродия рядовая | Облигатный сапротроф | Часто |
| Antrodia sinuos (Fr.) Karst. | Антродия извилистая | Облигатный сапротроф | Часто |
| Antrodia xantha (Fr.: Fr.) Ryvarden | Антродия золотистая | Облигатный сапротроф | Часто |
| Fomitopsis pinicola (Sw.: Fr.) Karst. | Окаймленный трутовик | Факультативный сапротроф | Часто |
| Fomitopsis rosea (Alb. et Schwein.: Fr) Karst. | Трутовик розовый | Облигатный сапротроф | Часто |
| Gloeophyllum odoratum (Wulfen: Fr.) Imaz. | Пахучий трутовик | Облигатный сапротроф | Часто |
| Gloeophyllum sepiarium (Wulfen: Fr.) Karst. | Трутовик заборный | Облигатный сапротроф | Часто |
| Leucogyrophana mollusca (Fr.) Pouzar | Леукогирофана мягкая | Облигатный сапротроф | Регулярно |
| Phellinus ferrugineofuscus (Karst.) Bourdot | Феллинус ржаво-бурый | Облигатный сапротроф | Регулярно |
| Phellinus nigrolimitatus (Romell) Bourd. et Galz. | Феллинус черноограниченный | Облигатный сапротроф | Регулярно |
| Phellinus viticola (Schwein. ex. Fr.) Donk. | Феллинус виноградный | Облигатный сапротроф | Регулярно |
| Phlebiopsis gigantea (Fr.: Fr.) Jülich | Флебиопсис гигантский | Облигатный сапротроф | Регулярно |
| Postia fragilis (Fr.) Jülich | Постия хрупкая | Облигатный сапротроф | Регулярно |
| Pycnoporellus fulgens (Fr.) Donk | Пикнопореллус блестящий | Облигатный сапротроф | Регулярно |
| Skeletocutis amorpha (Fr.) Kotl.et Pouzar | Скелетокутис бесформенный | Облигатный сапротроф | Регулярно |
| Skeletocutis odora (Sacc.) Ginns | Скелетокутис пахучий | Облигатный сапротроф | Регулярно |
| Trichaptum abietinum (Pers.: Fr.) Ryvarden | Трихаптум пихтовый | Облигатный сапротроф | Часто |
| Trichaptum fuscoviolaceum (Schmidt: Fr.) Kr. | Трихаптум буро-фиолетовый | Облигатный сапротроф | Регулярно |
| Trichaptum laricinum (Karst.) Ryvarden | Трихаптум лиственничный | Облигатный сапротроф | Регулярно |
Обозначения. Облигатный сапротроф использует для питания только мертвый субстрат; факультативный сапротроф и факультативный биотроф используют для питания как живой, так и мертвый субстрат. Встречаемость: часто — встречается на каждом пункте учета; регулярно — встречается не на каждом пункте учета.
Повторимся, в представленном списке грибов биотрофного и сапротрофного комплексов приведены виды, составляющие основу грибного дереворазрушающего сообщества еловых лесов южной тайги, которое в полном объеме включает значительно большее количество.
ЗАКЛЮЧЕНИЕ
Коренные разновозрастные ельники южной тайги имеют разное количество поколений в возрастных рядах древостоев с разными объемами деревьев в возрастных поколениях, что определяет их динамическое положение в сукцессионном ряду развития биогеоценоза.
Поставленные в один временной ряд все древесные фракции коренных ельников различных сукцессионных положений — от ретроспективных значений (–60 лет) и до перспективных значений предельного возраста деревьев первых поколений древостоев (+ ~320 лет) — позволили оценить в физических параметрах процессы трансформации фракций древесины в длительном временном пространстве с различной скоростью протекания процессов накопления и разложения древесной биомассы.
При этом в коренных девственных лесах сохраняется баланс воспроизводимой и разлагаемой древесины, и этот баланс служит одним из важнейших критериев устойчивого функционирования лесных сообществ. Это положение можно расценивать как важный факт функциональной структуры лесов.
Дереворазрушающие грибы сапротрофного комплекса «подстраивают» активность процесса разложения древесного отпада под динамику накопления древесины древостоем, сохраняя баланс накопления и разложения биомассы лесного сообщества.
В изучаемых ельниках корреляционная зависимость увеличения значений пораженности деревьев дереворазрушающими грибами с увеличением возраста древостоя очень высокая, почти функциональная.
СПИСОК ЛИТЕРАТУРЫ
Бондарцева М. А. Определитель грибов России. Афиллофоровые. СПб. Наука, 1998. Вып. 2. 391 с.
Гусев И. И. Строение и особенности таксации ельников Севера. М.: Лесн. пром-ть, 1964. 76 с.
Дыренков С. А. Структура и динамика таежных ельников. Л.: Наука, 1984. 176 с.
Исаев А. С., Суховольский В. Г., Хлебопрос Р. Г., Бузыкин А. И., Овчинникова Т. М. Моделирование лесообразовательного процесса: феноменологический подход / Мониторинг биологического разнообразия лесов России. М.: Наука. 2008. 451 с.
Курнаев C. Ф. Лесорастительное районирование СССР. М.: Наука, 1973. 203 с.
Морозов Г. Ф. О лесоводственных устоях / Избранные труды. М.: Лесная пром-ть, 1962. Т. 1. С. 459–474.
Правила санитарной безопасности в лесах (утв. приказом Министерства природных ресурсов и экологии РФ от 24 декабря 2013 г. № 613). 2013. 23 с.
Соловьев В. А. Микогенный ксилолиз, его экологическое и технологическое значение / Научные основы устойчивости лесов к дереворазрушающим грибам. М.: Наука, 1992. С. 140–171.
Стороженко В. Г. Датировка разложения валежника ели // Экология. 1990. № 6. С. 66–69.
Стороженко В. Г. Древесный отпад в коренных лесах Русской равнины. М.: КМК, 2011. 122 с.
Стороженко В. Г. Устойчивые лесные сообщества. Теория и эксперимент. М.: Гриф и К, 2007. 190 с.
Стороженко В. Г., Бондарцева М. А., Соловьев В. А., Крутов В. И. Научные основы устойчивости лесов к дереворазрушающим грибам. М.: Наука, 1992. 221 c.
Стороженко В. Г., Коткова В. М. Состояние коренных ельников и дереворазрушающие грибы (Basidiomycota) заповедника «Кологривский лес» (Костромской области) // Известия высших учебных заведений. Лесной журнал. 2013. № 3. С. 17–25.
Сукачев В. Н. Основы лесной биогеоценологии. М.: Наука, 1964. 458 с.
Третьяков Н. В., Горский П. В., Самойлович Г. Г. Справочник таксатора. М.-Л.: Гослесбумиздат, 1952. 853 с.
Niemelä T. Polypores of Finland and adjacent Russia // Norrlinia. 2001. No 8. P. 1–120.
Niemelä T. Käävät, puiden sienet. Polypores, lignicolous fungi // Norrlinia. 2005. No 13. P. 1–320.
Ryvarden L., Gilbertson R. L. European Polypores: Part. 1. Abortiporus-Lindtneria. Oslo: Fungiflora, 1993. P. 1–387.
Ryvarden L., Gilbertson R. L. European Polypores: Part. 2. Meripilus-Tyromyces. Oslo: Fungiflora, 1994. P. 388–743.
Tikkanen O.-P., Ruokolainen A., Heikkilä R. Recovery of boreal forest structures near abandoned villages in Western White Sea Karelia, Russia // Scandinavian Journal of Forest Research. 2014. Vol. 29. No 2. P. 152–161. DOI: 10.1080/02827581.2014.881543.
Рецензент: д. б. н. Евстигнеев О. И.





