- DOI: 10.31509/2658-607x-202581-162
- УДК 574.45
Распределение фитомассы и содержание азота и углерода в хвое сосны обыкновенной в средней тайге Республики Карелия
![]()
© 2025 г. Е. В. Новичонок*, Н. А. Галибина, К. М. Никерова
Институт леса КарНЦ РАН
Россия, Республика Карелия, 185910, Петрозаводск, ул. Пушкинская, 11
*E-mail: enovichonok@inbox.ru
Поступила в редакцию: 24.09.2024
После рецензирования: 17.02.2025
Принята к печати: 14.03.2025
Мониторинг фитомассы и продукции древесных растений является одной из приоритетных задач лесной экологии в связи с их ролью в глобальном балансе углерода. Выполнена оценка годичной продукции хвои, распределения фитомассы хвои по возрастам, а также содержания азота и углерода в хвое деревьев Pinus sylvestris в насаждениях разного возраста в разных лесорастительных условиях. Самые высокие значения годичной продукции хвои доминирующего элемента насаждения были отмечены в 180-190-летних сосняках черничных на песчаных и супесчаных подзолах. При этом годичная продукция хвои деревьев на вырубках была значимо ниже, чем в древостоях 40-50-, 80-100- и 180-190-лет. Доля фитомассы хвои разного возраста в общей фитомассе хвои различалась между молодыми деревьями на вырубке (до 10 лет) и деревьями большего возраста (40-50, 80-100 и 180-190-летние деревья). У молодых деревьев на вырубке доля хвои текущего года и однолетней хвои составляет почти 90%, у взрослых деревьев ˗ около 50% от всей массы хвои. Эти данные необходимо учитывать при построении моделей газообмена и потоков углерода в древостоях разного возраста в связи с зависимостью скорости фотосинтеза от возраста хвои. При увеличении возраста деревьев отмечалось более высокое содержание углерода в хвое. Обсуждается важность точной оценки содержания углерода в фитомассе для более корректной оценки запасов углерода в фитоценозе.
Ключевые слова: сосна обыкновенная, азот, углерод, доля фитомассы хвои, лесорастительные условия, сосняки разного возраста
Леса, в том числе бореальные, играют важную роль в глобальном балансе углерода и его депонировании (Nunes et al., 2020; Anderson-Teixeira et al., 2021). В связи с этим мониторинг их фитомассы и продукции является одной из приоритетных задач лесной экологии (Усольцев и др., 2015, 2022), а точная оценка аккумулированного углерода в различных компонентах лесных биоценозов позволит количественно оценить величину стока углерода из атмосферы и роль лесов в глобальном цикле углерода.
Фитомасса древесных растений и их продукция сильно варьируют в зависимости от возраста деревьев, а также от доступности питательных веществ, климатических и погодных условий (Portsmuth et al., 2005; Lim et al., 2017). Продуктивность растений в бореальных лесах ограничена недостатком доступного азота (Högberg et al., 2017). С учетом тесной связи между циклами азота и углерода (Tateno, Chapin, 1997) и зависимостью скорости роста и продуктивности растений от доступности азота (Portsmuth et al., 2005; Lim et al., 2017) необходима совместная оценка фитомассы и содержания азота и углерода в органах растений.
В настоящее время для оценки фитомассы и продукции лесных древостоев широко развиваются методы дистанционного зондирования (Rahimzadeh-Bajgiran et al., 2020; Labrière et al., 2023). Для их корректировки и повышения точности оценок необходимы прямые измерения содержания углерода в растительных тканях, а также фитомассы и продукции на уровне растительных сообществ и отдельных деревьев (Усольцев и др., 2016; Барталев, 2022; Tolunay, 2009; Labrière et al., 2023). Однако имеющихся данных об аккумулировании углерода в фитомассе и первичной продукции лесных биогеоценозов недостаточно для решения этих задач (Усольцев, Залесов, 2005; Вараксин и др., 2008).
Скорость протекания большинства физиологических процессов изменяется с увеличением возраста хвои. Скорость фотосинтеза зависит от возраста хвои (Teskey et al., 1984; Warren, 2006; Robakowski, Bielinis, 2017) и, таким образом, распределение фитомассы хвои по возрастам в кроне дерева оказывает непосредственное влияние на его продуктивность. Знание распределения фитомассы хвои разного возраста в кроне дерева и/или пологе насаждения имеет основополагающее значение для изучения основных обменных процессов на уровне древостоя (Li et al., 2006; Beeck et al., 2010). Эти данные необходимы для более точного моделирования и расчета общей скорости фотосинтеза отдельных деревьев и всего древостоя.
Цель исследования – оценить годичную продукцию хвои, распределение фитомассы хвои по возрастам, а также содержания азота и углерода в хвое деревьев Pinus sylvestris в насаждениях разного возраста и в разных лесорастительных условиях.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование проводили в средней подзоне тайги на северо-западе России (Республика Карелия). Климат в районе исследования относится к атлантико-арктической зоне умеренного пояса. Среднегодовая температура воздуха в районе исследования составляет около 3оС, среднегодовая сумма осадков – около 600 мм (Климат Карелии, 2004; Назарова, 2014).
Исследование проводили на растениях сосны, произрастающих на постоянных пробных площадях (ППП) тестового полигона интенсивного уровня «Кивач» (табл. 1) и на ППП (три типа БГЦ), заложенных в эксплуатационных лесах. 15 ППП трех типов БГЦ находились на трех стадиях восстановительной (послерубочной) сукцессии: (1) вырубки давностью до 10 лет, возникшие в результате рубки сосняков зеленомошных на песчаных подзолах; (2) средневозрастные (40-50 лет) сосняки зеленомошные на песчаных подзолах; (3) приспевающие (80-100 лет) сосняки зеленомошные естественного происхождения на песчаных подзолах (табл. 2). Тестовый полигон «Кивач» и сеть ППП в эксплуатационных лесах в зависимости от стадии послерубочной сукцессии созданы в 2023-2024 гг. сотрудниками Института леса КарНЦ РАН по проекту «Разработка системы мониторинга бюджета углерода в лесах Восточной Фенноскандии» в рамках реализации Важнейшего инновационного проекта государственного значения (ВИП ГЗ) «Единая национальная система мониторинга климатически активных веществ» в части разработки системы наземного и дистанционного мониторинга пулов углерода и потоков парниковых газов на территории Российской Федерации, создания системы учета данных о потоках климатически активных веществ и бюджете углерода в лесах и других наземных экологических системах.
Изучение влияния лесорастительных условий на годичную продукцию хвои, распределение фитомассы хвои по возрастам и содержание углерода и азота в хвое проводили на ППП тестового полигона интенсивного уровня «Кивач», объединенных в группы в соответствии с типом леса, возрастом древостоя, напочвенным покровом и типом почвы (табл. 1). Влияние возраста деревьев на исследуемые показатели изучали на примере сосняков зеленомошных на песчаных подзолах в ряду: вырубки до 10 лет (группа 1, ППП В-1…5) – средневозрастные сосняки (группа 2, ППП Ср-1…5) – приспевающие сосняки (группа 3, ППП Пр-1…6) – старовозрастные сосняки (группа 4, ППП I группы тестового полигона) (табл. 1, 2).
Таблица 1. Характеристика ППП тестового полигона «Кивач»
| Группа ППП | Число ППП, шт. | Тип леса | Тип почвы* | Доминирующий элемент леса, возраст, лет | Характеристика деревьев, с которых был отобран растительный материал | ||
| высота, м | диаметр на высоте 1.3 м, см | количество деревьев, шт. | |||||
| I | 5 | Сосняк черничный | Песчаные подзолы | Сосна, 180-190 | 26.2±0.7 | 33.3±1.1 | 8 |
| II | 4 | Сосняк черничный | Супесчаные подзолы | Сосна, 180-190 | 27.5±1.3 | 36.2±2.2 | 9 |
| IV | 2 | Сосняк брусничный | Песчаные подзолы | Сосна, 180 | 24.5±0.5 | 28.1±1.2 | 4 |
| VI | 3 | Сосняк черничный | Суглинистые на подзолистых почвах | Сосна, 140-180 | 28.4±0.9 | 44.3±4.7 | 4 |
| VII | 3 | Смешанные сосново-еловые черничные насаждения | Торфяно-глееземы | Сосна, 160-180 | 28.2±0.4 | 40.6±3.0 | 3 |
| VIII | 3 | Смешанные сосново-еловые древостои | Пелоземы | Сосна, 85-220 | 34.1±6.2 | 28.0±1.7 | 3 |
*Тип почвы был определен в соответствии с методическими рекомендациями, представленными в работе: Шишов и др., 2004.
Таблица 2. Таксационная характеристика насаждений на опытных объектах
| ППП | Состав | Доминирующий элемент насаждения | Характеристика деревьев, с которых был отобран растительный материал | |||||
| возраст, лет | густота, шт. га-1 | средняя высота, м | средний диаметр, см* | средняя высота, м | средний диаметр, см* | количество деревьев, шт. | ||
| Группа 1 – молодняки на вырубках | ||||||||
| В1 | 6.9Слк1.5С1.5Б0.1Е | 5 | 2520 | 0.6 | 0.92 | 0.60±0.01 | 0.90±0.05 | 3 |
| В2 | 5.5Слк3.1С1.4Б | 5 | 4160 | 0.6 | 0.71 | 0.59±0.02 | 0.76±0.05 | 3 |
| В3 | 5.5С3.1Слк1.4Б | 5 | 3440 | 0.3 | 0.65 | 0.42±0.01 | 0.67±0.08 | 3 |
| В4 | 4.7Слк2.8С2.1Б0.4Ос | 6 | 3000 | 1.0 | 1.20 | 0.92±0.01 | 1.25±0.15 | 3 |
| В5 | 3.9Б3.3Слк2.6С0.2Ос | 6 | 1440 | 1.0 | 1.18 | 1.02±0.02 | 1.40±0.16 | 3 |
| Группа 2 – средневозрастные древостои | ||||||||
| СР1 | 6.2Слк3.8С ед.Е ед.Б | 43 | 1204 | 13.2 | 12.3 | 13.25 | 8.9 | 1 |
| СР2 | 6.5Слк3.4С0.1Б | 43 | 1284 | 11.1 | 10.7 | 12.80 | 9.8 | 1 |
| СР3 | 5.5Слк4.4С0.1Б | 43 | 1052 | 12.3 | 12.1 | 11.80 | 9.6 | 1 |
| СР4 | 9.2Слк0.8С | 48 | 1120 | 12.5 | 14.7 | 17.17 | 12.5 | 1 |
| СР5 | 9.3Слк0.7С | 48 | 1180 | 14.8 | 15.2 | 14.14 | 13.5 | 1 |
| Группа 3 – приспевающие (спелые) древостои | ||||||||
| ПР1 | 7.2С952С1400.5Е0.2С0.1Б | 95 | 608 | 21.8 | 21.4 | 22.8±0.6 | 22.9±1.0 | 2 |
| ПР2 | 8.6С950.7Е0.6С0.1С ед.Б | 95 | 708 | 20.1 | 20.9 | 19.8±1.0 | 20.0±0.8 | 2 |
| ПР3 | 8.2С951.3С1400.4Е0.1С ед.Б | 95 | 760 | 20.3 | 20.0 | 21.6 | 19.8 | 1 |
| ПР4 | 9.6С950.2С1400.1Е0.1С ед.Б | 95 | 740 | 18.8 | 20.8 | 21.3±0.4 | 21.0±0.6 | 2 |
| ПР5 | 8.7С950.9С1400.2Е0.1Б0.1С | 95 | 768 | 21.3 | 20.7 | 19.3 | 21.6 | 2 |
*Для молодняков на вырубках указан диаметр на уровне ½ от высоты подроста, для деревьев средневозрастных и приспевающих древостоев – диаметр на высоте 1.3 м.
Растительный материал отбирали с 31 июля по 14 августа 2023 г. (тестовый полигон «Кивач») и с 25 июля по 14 августа 2024 г. (тестовый полигон «Кивач», сосновые насаждения разного возраста (группа 1, 2, 3)). В этот период ростовые процессы уже закончены, а массовое пожелтение и опадение хвои еще не началось (Кищенко, 2020 а, б).
На всех ППП на основании результатов таксационных исследований были выбраны модельные деревья, которые соответствовали средним значениям по диаметру на высоте 1.3 м (или на уровне ½ от высоты подроста для молодняков на вырубках), рассчитанным для каждой ППП, а также не имели видимых повреждений ствола/кроны, признаков потери верхушечного побега и признаков снижения приростов. Характеристика модельных деревьев и количество отобранных деревьев на каждой ППП представлены в табл. 1 и 2.
С модельных деревьев полигона «Кивач» и деревьев из приспевающих древостоев (группа 3) из средней трети кроны с южной стороны было отобрано по одной незатененной ветви. Ветви были разделены на побеги, согласно возрасту хвои. В лабораторных условиях определяли массу хвои для всех лет жизни, рассчитывали долю хвои каждого возраста, а также величину годичной продукции хвои дерева с использованием аллометрического уравнения (Усольцев и др., 2016; Методические рекомендации …, 2023). В средневозрастных древостоях (группа 2) было спилено пять модельных деревьев. У деревьев обрубали и взвешивали все ветви с учетом их расположения в кроне дерева (из верхней, средней и нижней частей кроны). Затем отбирали по две модельные ветви из каждой части кроны. У модельных ветвей отделяли хвою с учетом возраста и после высушивания определяли фитомассу ветвей и хвои разных лет жизни. Полученные данные были использованы для расчета фитомассы хвои всего дерева, годичной продукции хвои дерева и доли хвои каждого возраста (Базилевич и др., 1978; Яковлев, 2024). В молодняках на вырубках (группа 1) выкапывали модельные деревья. Затем ветви были разделены на побеги согласно возрасту хвои. В лабораторных условиях определяли фитомассу хвои для всех лет жизни. На основе полученных данных была рассчитана фитомасса хвои всего дерева, годичная продукция хвои и доля хвои каждого возраста.
Годичную продукцию хвои (кг га-1) доминирующего элемента рассчитывали на основе данных о густоте доминирующего элемента и годичной продукции хвои в расчете на дерево.
Содержание углерода и азота было определено на элементном анализаторе Unicube Elementar (Elementar GmbH, Германия), калиброванного сертифицированным стандартом ацетанилида (200007435 Acetanilide Elementar Analysen systeme), с использованием 4-5 мг сухого растительного материала, высушенного до воздушно сухого состояния при 40оС и затем измельченного на лабораторной мельнице до гомогенного состояния «пудры».
Все pезультаты представлены в расчете на абсолютно сухую фитомассу.
Все анализы были проведены с использованием оборудования Центра коллективного пользования КарНЦ РАН (ИЛ КарНЦ РАН).
Статистическая обработка полученных данных была проведена с использованием программы STATISTICA (data analysis software system, version 10, StatSoft, Inc., 2011). Для определения влияния лесорастительных условий, возраста деревьев и возраста хвои на изученные показатели был проведен однофакторный дисперсионный анализ. Сравнение средних проводили, используя тест Бонферрони. Статистически значимыми считали различия при р<0.05. В таблицах указаны средние значения ± стандартная ошибка.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Годичная продукция хвои в разных лесорастительных условиях. У 180-190-летних деревьев сосны годичная продукция хвои изменялась в зависимости от условий произрастания (рис. 1). Максимальные значения годичной продукции хвои в расчете на дерево были отмечены в сосняке черничном на суглинистых подзолистых почвах (группа VI), минимальные – в сосняке брусничном на песчаных подзолах (группа IV) (рис. 1 A). Скорость роста и интенсивность формирования надземной фитомассы выше при лучших почвенных условиях (Vanninen et al., 1996; Wirth et al., 2002; Кищенко, 2019). В сосняке брусничном также отмечались одни из самых низких значений годичной фитомассы хвои доминирующего элемента (рис. 1 Б). Однако из-за высокой степени вариации между насаждениями разных групп ППП статистически значимых отличий не отмечалось. При этом самые высокие значения годичной продукции хвои доминирующего элемента были отмечены в сосняках черничных на песчаных и супесчаных подзолах. Сходные данные по продуктивности были получены и для средневозрастных древостоев в разных типах леса в Карелии, где было показано, что в сосняке брусничном годичный прирост фитомассы хвои на 20% ниже по сравнению с сосняком черничным (Кищенко, 2019).
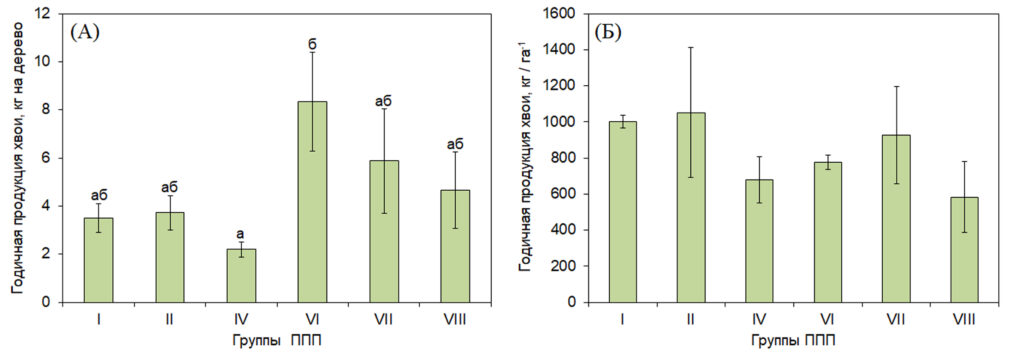
Рисунок 1. Годичная продукция хвои в расчете на дерево (А) и доминирующего элемента (Б) в группах ППП тестового полигона «Кивач»
Примечание: Характеристика групп ППП представлена в табл. 1. Разные строчные буквы обозначают достоверность отличий между группами ППП.
Годичная продукция хвои в расчете на дерево повышалась с увеличением возраста деревьев (рис. 2 А). Однако годичная продукция хвои доминирующего элемента была сходной в насаждениях 40-50, 80-100 и 180-190 лет, что связано с увеличением густоты насаждений со снижением их возраста. Годичная продукция хвои доминирующего элемента на вырубках (группа 1) была значимо ниже по сравнению с этим показателем для насаждений большего возраста (средневозрастные, приспевающие и старовозрастные сосняки) (рис. 2 Б).
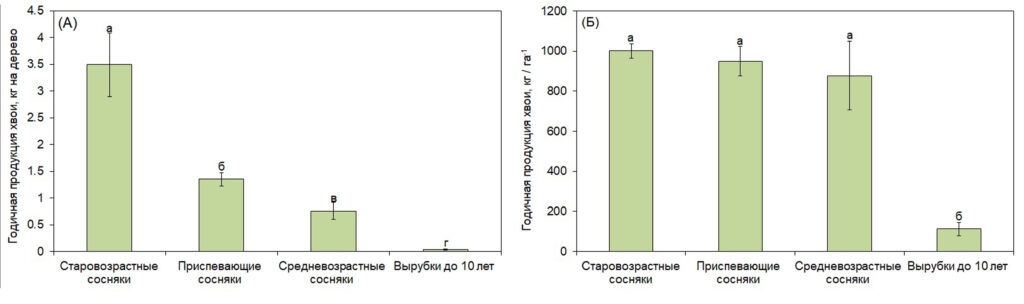
Рисунок 2. Годичная продукция хвои в расчете на дерево (А) и доминирующего элемента (Б) в древостоях разного возраста
Примечание: Характеристика насаждений разного возраста представлена в табл. 2. Разные строчные буквы обозначают достоверность отличий между насаждениями разного возраста.
Продолжительность жизни хвои и доля хвои разного возраста. У 180-190-летних и 80-100-летних деревьев сосны хвоя сохраняется 4-6 лет. У более молодых растений продолжительность жизни хвои меньше, и она составляет 5 лет у 40-50-летних растений и 3-4 года у 7-летних растений. Ранее было показано, что продолжительность жизни хвои видов рода Pinus увеличивается с увеличением возраста деревьев, а также зависит от климатических факторов и лесорастительных условий, в частности доступности питательных веществ и воды (Бабич и др., 2004; Тюкавина, Корепин, 2023; Pensa, Sellin, 2002; Xiao, 2003;). Однако мы не обнаружили разницу в продолжительности жизни хвои у 180-190-летних деревьев в разных лесорастительных условиях на ППП тестового полигона «Кивач». У большинства деревьев (58%) хвоя сохранялась в течение 5 лет, у 32% – 4 года и только у 10% деревьев хвоя сохранялась в течение 6 лет. Отсутствие разницы в продолжительности жизни хвои в разных лесорастительных условиях может быть связано с тем, что они в большей степени могут оказывать влияние на сохранность хвои, тогда как количество возрастных классов может оставаться постоянным (Pensa, Sellin, 2002). Cтабильность средней продолжительности жизни хвои при изменении типа леса может указывать на оптимальное значение данного показателя для конкретной возрастной группы (Тюкавина, Корепин, 2023). Изменение продолжительности жизни хвои влияет на соотношение ее фитомассы в кроне дерева и соответственно на скорость фотосинтеза и продукционный процесс. Доля фитомассы хвои разного возраста в общей фитомассе хвои на дерево различалась между молодыми деревьями (группа 1) и деревьями большего возраста (группы 2, 3, 4) (табл. 3). Между деревьями 40-50, 80-100 и 180-190 лет не было отмечено различий в доле фитомассы хвои разного возраста. Наиболее стабильным показателем была доля фитомассы однолетней хвои: у деревьев всех возрастных групп значение этого показателя было сходным. У молодых деревьев (группа 1) доля фитомассы хвои каждого возраста уменьшалась с увеличением возраста хвои, и максимальная фитомасса была отмечена для хвои текущего года, тогда как для более взрослых деревьев (группы 2-4) максимальная фитомасса была отмечена для однолетней и двухлетней хвои (табл. 3).
Таблица 3. Доля фитомассы хвои разного возраста (%) в общей фитомассе хвои в сосновых насаждениях разного возраста
| Возраст хвои | Молодняки на вырубках (группа 1) | Средневозрастные древостои 40-50 лет (группа 2) | Приспевающие (спелые) древостои 80-100 лет (группа 3) | 180-190-летний сосняк черничный (группа 4) |
| Текущий год | 61.7±4.7 а | 22.8±1.3 б | 24.6±1.8 б | 21.0±2.5 б |
| 1 год | 28.0±1.5 а | 27.2±1.2 а | 31.2±2.6 а | 30.3±2.2 а |
| 2 года | 11.7±4.9 а | 29.3±0.8 б | 29.5±1.8 б | 30.1±1.8б |
| 3 года | 2.3±2.1 а | 19.5±1.6 б | 13.8±1.6 б | 18.1±1.8б |
| 4 года | – | 2.7±1.3 а | 2.2±0.4 а | 5.4±0.9а |
| 5 лет | – | – | – | 7.0 |
Примечания: Прочерк – хвоя данного возрастного класса отсутствует. Разные строчные буквы обозначают достоверность отличий в доле фитомассы хвои разного возраста между насаждениями разного возраста. Характеристика насаждений разного возраста представлена в табл. 2.
Не было отмечено статистически значимых отличий в доле фитомассы хвои разного возраста в общей фитомассе хвои между 180-190-летними деревьями, произрастающими в разных лесорастительных условиях (табл. 4). Однако отмечалась тенденция более высокой доли однолетней хвои и снижения доли трехлетней хвои у деревьев сосны в смешанных сосново-еловых древостоях (ППП VIII) (табл. 4). Распределение фитомассы хвои сильно варьирует от размера и возраста деревьев, а также от доступности питательных веществ и климатических условий (Portsmuth et al., 2005; Li et al., 2006). У сосны обыкновенной в разных условиях доля хвои текущего года может варьировать в широких пределах (Xiao et al., 2003; Li et al., 2006; Tarvainen et al., 2016). Полученные нами данные согласуются с данными, полученными для сосны обыкновенной в Карелии и близких к ней регионах. Доля хвои текущего года в Карелии составляет 25%, в европейской части России колеблется от 26.5 до 31.6% от всей ее фитомассы (Иванчиков и др., 1982; Бабич и др., 2004).
Таблица 4. Доля фитомассы хвои разного возраста (%) в общей фитомассе хвои у 180-190-летних растений в разных лесорастительных условиях
| Возраст хвои | Группа ППП I | Группа ППП II | Группа ППП IV | Группа ППП VI | Группа ППП VII | Группа ППП VIII |
| Текущий год | 21.0±2.5 | 18.7±2.0 | 19.6±1.4 | 23.8±2.6 | 24.9±4.5 | 26.1±2.6 |
| 1 год | 30.3±2.2 | 38.1±3.1 | 37.7±6.0 | 36.1±2.7 | 29.4±3.3 | 41.2±2.7 |
| 2 года | 30.1±1.8 | 27.2±1.6 | 27.9±1.9 | 28.1±3.5 | 31.9±2.3 | 24.0±2.0 |
| 3 года | 18.1±1.8 | 14.8±2.0 | 15.4±2.8 | 12.4±2.9 | 13.2±3.6 | 8.7±0.9 |
| 4 года | 5.4±0.9 | 5.2 | 4.4±2.9 | 5.3±2.6 | 1.0±0.1 | – |
| 5 лет | 7.0 | – | 2.0 | – | – | – |
Примечания: Прочерк – хвоя данного возрастного класса отсутствует. Характеристика групп ППП представлена в табл. 1.
Скорость фотосинтеза снижается с увеличением возраста хвои (Teskey et al., 1984; Warren, 2006; Robakowski, Bielinis, 2017). Хвоя первых двух лет жизни имеет наибольшую скорость фотосинтеза (Teskey et al., 1984; Warren, 2006). У молодых деревьев на вырубке доля однолетней хвои и хвои текущего года жизни составляет почти 90%, у более взрослых деревьев (группа 2, 3, 4) ˗ около 50% от всей фитомассы хвои (табл. 3). Полученные различия необходимо учитывать при построении моделей газообмена и потоков углерода в древостоях разного возраста. Упущение возрастных изменений фотосинтетической способности могут привести к значительно менее точным оценкам газообмена в пологе хвойных деревьев (Ogée et al., 2003).
Содержание азота и углерода в хвое. Увеличение продолжительности жизни хвои отражает более высокую эффективность использования ресурсов и позволяет сохранять питательные вещества в менее благоприятных условиях произрастания (Pensa, Sellin, 2002; Li et al., 2006). Содержание азота в хвое было сходным в разных лесорастительных условиях (рис. 3 A). Мы отмечали тенденцию к снижению содержания азота в хвое с увеличением возраста хвои, вызванное ретранслокацией азота из стареющей и опадающей хвои, в том числе в оставшуюся на дереве хвою (Field, 1983; Helmisaari, 1992). У молодых деревьев на вырубке содержание азота в двухлетней и трех-четырех-летней хвое было статистически ниже по сравнению с его содержанием у деревьев других возрастных групп (рис. 3 Б), что связано с более интенсивной его ретранслокацией в связи с низкой сохранностью хвои этих возрастов у деревьев этой возрастной группы (табл. 3). Отмеченное нами содержание азота было в два и более раз ниже оптимального (1.88-2.2%), установленного для саженцев сосны обыкновенной в лабораторных условиях (Ingestad, 1979; Ingestad, Kӓhr, 1985). Для более взрослых деревьев сосны (20 лет и старше) было показано, что оптимальное содержание азота в хвое текущего года, при котором отмечается максимальная скорость роста, составляет 14-15 мг г-1 (1.4-1.5%) (Sikstrӧm et al., 1998).
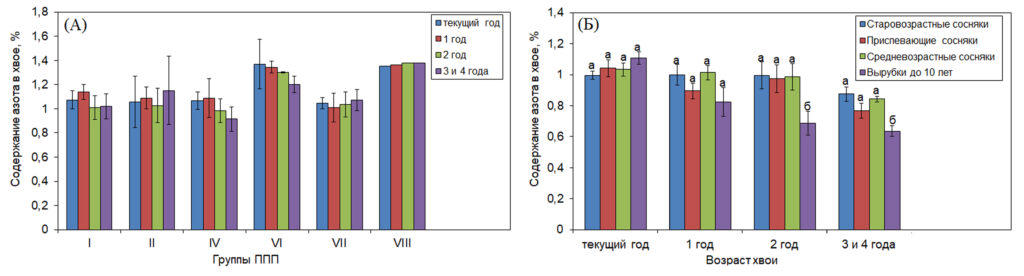
Рисунок 3. Содержание азота в хвое разных лет жизни у деревьев в группах ППП тестового полигона «Кивач» (А) и в сосновых насаждениях разного возраста (Б)
Примечания: Разные строчные буквы обозначают достоверность отличий в содержании азота в хвое у деревьев, произрастающих в сосновых насаждениях разного возраста отдельно для каждого возраста хвои. Характеристика групп ППП и насаждений разного возраста представлена в табл. 1 и 2.» width=»1024″ height=»276″ /> Рисунок 3. Содержание азота в хвое разных лет жизни у деревьев в группах ППП тестового полигона «Кивач» (А) и в сосновых насаждениях разного возраста (Б)
Примечания: Разные строчные буквы обозначают достоверность отличий в содержании азота в хвое у деревьев, произрастающих в сосновых насаждениях разного возраста отдельно для каждого возраста хвои. Характеристика групп ППП и насаждений разного возраста представлена в табл. 1 и 2.
Для более точного расчета количества углерода, аккумулированного в фитомассе, необходимы знания об изменении его содержания в хвое разного возраста, а также при изменении возраста древостоя. При расчете запасов углерода в наземной фитомассе исходят из того, что содержание углерода в хвое сосны обыкновенной составляет 45% (Бабич и др., 2010; Сунгурова и др., 2019; Клевцов, Тюкавина, 2023), а среднее содержание углерода в пересчете на дерево ˗ 50% (Pohjola, Valsta, 2007; Tolunay, 2009). Мы получили, что содержание углерода в хвое разных лет жизни у деревьев разного возраста выше 45 % и колеблется в диапазоне 49.7 ˗ 55.8%, а у отдельных деревьев достигает 65.4% (рис. 4). В работах также было отмечено, что использование коэффициента 0.45 (из расчета, что доля углерода в хвое составляет 45%) при оценке запаса углерода в фитомассе хвои занижает реальные значения и может привести к недооценке аккумулированного углерода (Tolunay, 2009). Точная оценка углерода в хвое важна и потому, что в сосновых древостоях доля хвои в опаде велика (Albrektson, 1988; Pausas, 1997), а его содержание в подстилке может оказывать влияние на скорость ее разложения (Vestgarden, 2001) и, следовательно, на почвообразовательные процессы и выделение углерода в процессе почвенного дыхания.
Концентрация углерода в хвое сосны обыкновенной варьирует в различных почвенно-климатических условиях (Сухарева и др., 2023; Janssens et al., 1999; Węgiel, Polowy, 2020). В нашем исследовании содержание углерода у деревьев в разных лесорастительных условиях отличалось только у однолетней хвои (рис. 4 А), и было сходным у деревьев одного возраста в разновозрастной хвое (рис. 4 Б). Сходное содержание углерода в разновозрастной хвое может быть связано с перераспределением углерода между различными органами в пользу корней, побегов и ствола, в которых ассимилированный углерод депонируется (Сухарева и др., 2023; Bert, Danjon, 2006; Tolunay, 2009). Однако отмечалось увеличение его содержания с увеличением возраста деревьев (рис. 4 Б).
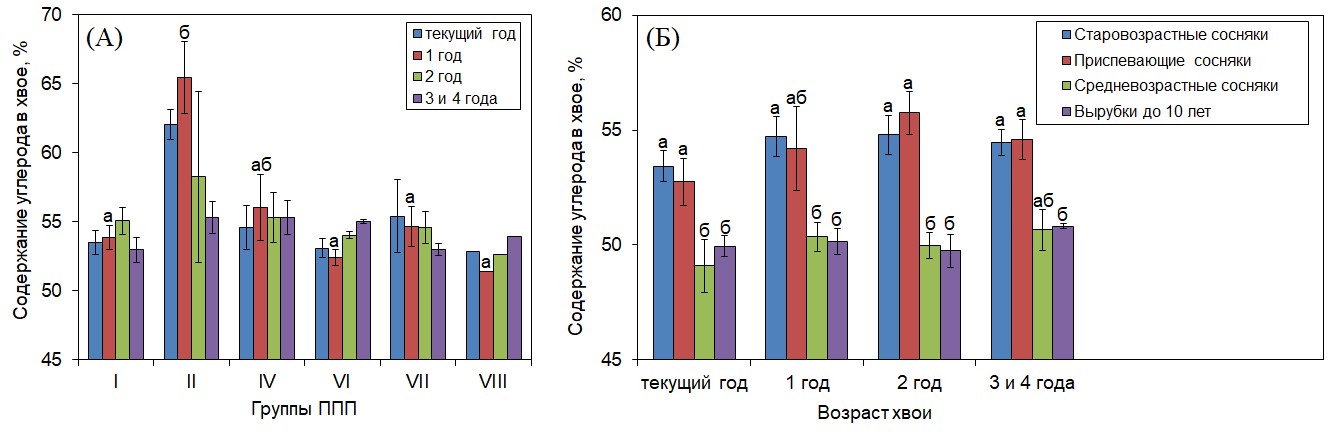
Рисунок 4. Содержание углерода в хвое разных лет жизни у деревьев в группах ППП тестового полигона «Кивач» (А) и в сосновых насаждениях разного возраста (Б)
Примечания: Разные строчные буквы обозначают достоверность отличий в содержании углерода в хвое у деревьев разных групп ППП тестового полигона «Кивач» (А) и произрастающих в сосновых насаждениях разного возраста отдельно для каждого возраста хвои (Б). Характеристика групп ППП и насаждений разного возраста представлена в табл. 1 и 2.
ЗАКЛЮЧЕНИЕ
Проведенное исследование показало, что лесорастительные условия оказывают влияние на годичную продукцию хвои, а также на содержание углерода в однолетней хвое. У более взрослых деревьев увеличиваются годичная продукция хвои и содержание углерода в хвое, но снижается доля хвои первых двух лет жизни. Полученные данные по изменению содержания годичной продукции хвои и содержанию углерода при изменении возраста сосновых насаждений и лесорастительных условий необходимо учитывать при оценке запасов фитомассы и углерода, а также депонирующей способности сосновых насаждений бореальной зоны. Полученное нами значение содержания углерода в хвое (49.7 ˗ 55.8%) значительно выше общепринятого для оценки запасов углерода в фитомассе (45%), что приводит к недооценке запасов углерода в сосновых насаждениях средней подзоны тайги.
БЛАГОДАРНОСТИ
Работа выполнена в рамках реализации важнейшего инновационного проекта государственного значения «Разработка системы наземного и дистанционного мониторинга пулов углерода и потоков парниковых газов на территории Российской Федерации, обеспечение создания системы учета данных о потоках климатически активных веществ и бюджете углерода в лесах и других наземных экологических системах» (рег. № 123030300031-6).
Авторы благодарят сотрудников ИЛ КарНЦ РАН за проведение геоботанических и лесотаксационных описаний пробных площадей и подбор объектов для исследования, а также сотрудников аналитической лаборатории ИЛ КарНЦ РАН за проведение анализов по определению содержания углерода и азота в хвое.
СПИСОК ЛИТЕРАТУРЫ
Бабич Н. А., Клевцов Д. Н., Евдокимов И. В. Зональные закономерности изменения фитомассы культур сосны: монография. Архангельск: САФУ, 2010. 140 с.
Бабич Н. А., Мерзленко М. Д., Евдокимов И. В. Фитомасса культур сосны и ели в европейской части России. Архангельск, 2004. 112 с.
Базилевич Н. И., Титлянова А. А., Смирнов В. В., Родин Л. Е., Нечаева Н. Т., Левин Ф. И. Методы изучения биологического круговорота в различных природных зонах: монография. М.: Мысль,1978. 184 с.
Барталев С. А. Методология комплексного использования спутниковых данных дистанционного зондирования, выборочных наземных наблюдений и моделирования для мониторинга бюджета углерода в лесах России (лекция) // Материалы 20-й Международной конференции «Современные проблемы дистанционного зондирования Земли из космоса». Москва, 14˗18 ноября 2022. М.: ИКИ РАН, 2022. C. 499. DOI: 10.21046/20DZZconf-2022a
Вараксин Г. С., Поляков В. И., Люминарская М. А. Биологическая продуктивность сосны обыкновенной в средней Сибири // Лесоведение. 2008. № 3. С. 14–19.
Иванчиков А. А., Зябченко С. С., Софронова Г. И. Ресурсы сосновой зелени в лесах Карелии // Комплексное ведение хозяйства в сосновых лесах: тезисы докладов. Гомель, 1982. С. 128–130.
Кищенко И. Т. Сезонное формирование надземной фитомассы средневозрастных сосновых древостоев различных типов леса в средней тайге // Лесоведение. 2019. № 1. С. 19–28.
Кищенко И. Т. Влияние климатических факторов на сезонное развитие хвойных лесообразующих видов в таежной зоне (Карелия) // Известия высших учебных заведений. Лесной журнал. 2020a. №3. С. 72–82.
Кищенко И. Т. Cезонный рост вегетативных органов Pinus sylvestris L. в разных типах лесных сообществ таежной зоны (Kарелия) // Norwegian Journal of Development of the International Science. 2020б. № 43. С. 13–17.
Клевцов Д. Н., Тюкавина О. Н. Запасы углерода в 40-летних культурах сосны обыкновенной // Известия высших учебных заведений. Лесной журнал. 2023. № 5 (395). С. 195–203.
Климат Карелии: Изменчивость и влияние на водные объекты и водосборы / под ред. Н. Н. Филатова. Петрозаводск: КарНЦ РАН, 2004. 224 с.
Методические рекомендации по отбору и камеральной обработке растительных образцов для оценки динамики запасов углерода в лесных, степных и тундровых экосистемах в части разработки системы наземного и дистанционного мониторинга пулов углерода и потоков парниковых газов на территории Российской Федерации, создания системы учета данных о потоках климатически активных веществ и бюджете углерода в лесах и других наземных экологических системах: версия 2023-1 / Н. В. Лукина, М. А. Данилова, Е. А. Иванова и др. Москва, 2023. 27 с.
Назарова Л. Е. Климат Республики Карелия (Россия): температура воздуха, изменчивость и изменения // Геополитика и экогеодинамика регионов. 2014. Т. 10. № 1. C. 746–749.
Сунгурова Н. Р., Худяков В. В., Страздаускас С. Е. Сравнительная структура углеродного пула в надземной фитомассе культур сосны и ели // Известия высших учебных заведений. Лесной журнал. 2019. № 3. С. 159–165.
Сухарева Т. А., Иванова Е. А., Ершов В. В., Зенкова И. В., Корнейкова М. А., Штабровская И. М., Сошина А. С. Содержание и запасы углерода и азота в наземных экосистемах Мурманской области // Вопросы лесной науки. 2023. Т. 6. № 2. Статья № 125. DOI: 10.31509/2658-607x-202362-125
Тюкавина О. Н., Корепин Д. Ю. Продолжительность жизни хвои сосны обыкновенной (Pinus sylvestris L.) // Resources and Technology. 2023. Vol. 20. № 3. P. 48–59.
Усольцев В. А., Залесов С. В. Методы определения биологической продуктивности насаждений. Екатеринбург: УГЛТУ, 2005. 147 с.
Усольцев В. А., Субботин К. С., Часовских В. П. Изменение удельной первичной продукции сосновых насаждений по трансконтинентальным климатическим градиентам Eвразии // Эко-ПОТЕНЦИАЛ. 2015. № 3 (11). С. 24–31.
Усольцев В. А., Цепордей И. С., Данилин И. М. Разработка модели биомассы Picea L. и Abies L. для региональных условий Евразии // Известия высших учебных заведений. Лесной журнал. 2022. № 6. С. 38–54.
Усольцев В. А., Часовских В. П., Норицина Ю. В., Норицин Д. В. Аллометрические модели фитомассы деревьев для лазерного зондирования и наземной таксации углеродного пула в лесах Евразии: сравнительный анализ // Сибирский лесной журнал. 2016. № 4. С. 68–76.
Шишов Л. Л., Тонконогов В. Д., Лебедева И. И., Герасимова М. И. Классификация и диагностика почв России. Смоленск: Oйкумена, 2004. 342 с.
Яковлев А. А. Влияние почвенных условий на формирование растительных сообществ на постагрогенных и лесных землях (на примере Ленинградской области): Дисс. … к.с-х.н. (специальность: 4.1.6). Санкт-Петербург, 2024. 354 c.
Albrektson A. Needle litterfall in stands of Pinus sylvestris L. in Sweden, in relation to site quality, stand age and latitude // Scandinavian Journal of Forest Research. 1988. Vol. 3. Issue 1–4. P. 333–342.
Anderson-Teixeira K. J., Herrmann V., Morgan R. B., Bond-Lamberty B., Cook-Patton S. C., … & Wang M. M. Carbon cycling in mature and regrowth forests globally // Environmental Research Letters. 2021. Vol. 16. No 5. Article number 053009. DOI:10.1088/1748-9326/abed01
Beeck M. O., Gielen B., Jonckheere I., Samson R., Janssens I. A., Ceulemans R. Needle age-related and seasonal photosynthetic capacity variation is negligible for modelling yearly gas exchange of a sparse temperate Scots pine forest // Biogeosciences. 2010. Vol. 7. No 1. P. 199–215. DOI: 10.5194/bg-7-199-2010
Bert D., Danjon F. Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.) // Forest Ecology and Management. 2006. Vol. 222. Issue 1–3. P. 279–295.
Field C. Allocating leaf nitrogen for the maximization of carbon gain: leaf age as a control on the allocation program // Oecologia. 1983. Vol. 56. P. 341–347.
Helmisaari H.-S. Nutrient retranslocation within the foliage of Pinus sylvestris // Tree Physiology. 1992. Vol. 10. No 1. P. 45–58.
Högberg P., Näsholm T., Franklin O., Högberg M. N. Tamm Review: On the nature of the nitrogen limitation to plant growth in Fennoscandian boreal forests // Forest Ecology and Management. 2017. Vol. 403. P. 161–185.
Ingestad T. Mineral nutrient requirements of Pinus sylvestris and Picea abies seedlings // Physiologia Plantarum. 1979. Vol. 45. No 4. P. 373–380.
Ingestad T., Kӓhr M. Nutrition and growth of coniferous seedlings at varied relative nitrogen addition rate // Physiologia Plantarum. 1985. Vol. 65. No 2. P. 109–116.
Janssens I. A., Sampson D. A., Cermak J., Meiresonne L., Riguzzi F., Overloop S., Ceulemans R. Above- and belowground phytomass and carbon storage in a Belgian Scots pine stand // Annals of Forest Science. 1999. Vol. 56. No 2. P. 81–90.
Labrière N., Davies S. J., Disney M. I., Duncanson L. I., Herold M., Lewis S. L., … & Chave J. Toward a forest biomass reference measurement system for remote sensing applications // Global Change Biology. 2023. Vol. 29. No 3. P. 827–840. DOI: 10.1111/gcb.16497
Li M. H., Kräuchi N., Dobbertin M. Biomass distribution of different-aged needles in young and old Pinus cembra trees at highland and lowland sites // Trees. 2006. Vol. 20. P. 611–618.
Lim H., Oren R., Linder S., From F., Nordin A., Fahlvik N., Lundmark T., Näsholm T. Annual climate variation modifies nitrogen induced carbon accumulation of Pinus sylvestris forests // Ecological Applications. 2017. Vol. 27. No 6. P. 1838–1851. DOI: 10.1002/eap.1571
Nunes L. J. R., Meireles C. I. R., Pinto Gomes C. J., Almeida Ribeiro N. M. C. Forest contribution to climate change mitigation: management oriented to carbon capture and storage // Climate. 2020. Vol. 8. No 2. Article number 21. DOI: 10.3390/cli8020021
Ogée J., Brunet Y., Loustau D., Berbigier P., Delzon S. MuSICA, a CO2, water and energy multilayer, multileaf pine forest model: evaluation from hourly to yearly time scales and sensitivity analysis // Global Change Biology. 2003. Vol. 9. No 5. P. 697–717. DOI: 10.1046/j.1365-2486.2003.00628.x
Pausas J. G. Litter fall and litter decomposition in Pinus sylvestris forests of the eastern Pyrenees // Journal of Vegetation Science. 1997. Vol. 8. No 5. P. 643–650.
Pensa M., Sellin A. Needle longevity of Scots pine in relation to foliar nitrogen content, specific leaf area, and shoot growth in different forest types // Canadian Journal of Forest Research. 2002. Vol. 32. No 7. P. 1225–1231.
Pohjola J., Valsta L. Carbon credits and management of Scots pine and Norway spruce stands in Finland // Forest Policy and Economics. 2007. Vol. 9. No 7. P. 789–798.
Portsmuth A., Niinemets Ü., Truus L., Pensa M. Biomass allocation and growth rates in Pinus sylvestris are interactively modified by nitrogen and phosphorus availabilities and by tree size and age // Canadian Journal of Forest Research. 2005. Vol. 35. No 10. P. 2346–2359.
Rahimzadeh-Bajgiran P., Hennigar C., Weiskittel A., Lamb S. Forest potential productivity mapping by linking remote-sensing-derived metrics to site variables // Remote Sensing. 2020. Vol. 12. No 12. Article number 2056. DOI: 10.3390/rs12122056
Robakowski P., Bielinis E. Needle age dependence of photosynthesis along a light gradient within an Abies alba crown // Acta Physiologiae Plantarum. 2017. Vol. 39. Article number 83. DOI: 10.1007/s11738-017-2376-y
Sikström U., Nohrstedt H.-Ö., Pettersson F., Jacobson S. Stem-growth response of Pinus sylvestris and Picea abies to nitrogen fertilization as related to needle nitrogen concentration // Trees. 1998. Vol. 12. P. 208–214.
Tarvainen L., Lutz M., Räntfors M., Näsholm T., Wallin G. Increased needle nitrogen contents did not improve shoot photosynthetic performance of mature nitrogen-poor Scots pine trees // Frontiers in Plant Science. 2016. Vol. 7. Article number 1051. DOI: 10.3389/fpls.2016.01051
Tateno M., Chapin F. S. The logic of carbon and nitrogen interactions in terrestrial ecosystems // The American Naturalist. 1997. Vol. 149. No 4. P. 723–744.
Teskey R. O., Grier C. C., Hinckley T. M. Change in photosynthesis and water relations with age and season in Abies amabilis // Canadian Journal of Forest Research. 1984. Vol. 4. No 1. P. 77–84.
Tolunay D. Carbon concentrations of tree components, forest floor and understorey in young Pinus sylvestris stands in north-western Turkey // Scandinavian Journal of Forest Research. 2009. Vol. 24. No 5. P. 394–402.
Vanninen P., Ylitalo H., Sievänen R., Mäkelä A. Effects of age and site quality on the distribution of biomass in Scots pine (Pinus sylvestris L.) // Trees. 2006. Vol. 10. P. 231–238. DOI: 10.1007/BF02185674
Vestgarden L. S. Carbon and nitrogen turnover in the early stage of Scots pine (Pinus sylvestris L.) needle litter decomposition: effects of internal and external nitrogen // Soil Biology and Biochemistry. 2001. Vol. 33. Issues 4–5. P. 465–474.
Warren C. R. Why does photosynthesis decrease with needle age in Pinus pinaster? // Trees. 2006. Vol. 20. P. 157–164.
Węgiel A., Polowy K. Aboveground carbon content and storage in mature Scots pine stands of different densities // Forests. 2020. Vol. 11. No 2. Article number 240. DOI: 10.3390/f11020240
Wirth С., Schulze E.-D., Kusznetova V., Milyukova I., Hardes G., Schulze B., Siry M., Vygodskaya N. Comparing the influence of site quality, stand age, fire and climate on aboveground tree production in Siberian Scots pine forests // Tree Physiology. 2002. Vol. 22. No 8. P. 537–552. DOI: 10.1093/treephys/22.8.537
Xiao C.-W., Yuste J. C., Janssens I. A., Roskams P., Nachtergale L., Carrara A., Sanchez B. Y., Ceulemans R. Above- and belowground biomass and net primary production in a 73-year-old Scots pine forest // Tree Physiology. 2003. Vol. 23. No 8. P. 505–516. DOI: 10.1093/treephys/23.8.505
Xiao Y. Variation in needle longevity of Pinus tabulaeformis forests at different geographic scales // Tree Physiology. 2003. Vol. 23. No 7. P. 463–471.
Рецензент: к.б.н., зав. лабораторией Сухарева Т. А.





