- DOI: 10.31509/2658-607x-202582-164
- УДК 581.143.28: 574.36(21)
Методические особенности учета продуктивности гипоарктических кустарников на полуострове Таймыр
![]()
© 2025 А. И. Бондарев1,2*, Л. В. Мухортова1, Д. А. Машуков1
1Институт леса им. В. Н. Сукачева Федерального исследовательского центра «Красноярский научный центр Сибирского отделения Российской академии наук»
Россия, 660036 Красноярск, Академгородок, 50/28
2Центр по проблемам экологии и продуктивности лесов им. А. С. Исаева Российской академии наук
Россия, 117997 Москва, ул. Профсоюзная, 84/32, стр. 14
*E-mail: abond822@yandex.ru
Поступила в редакцию: 22.11.2024
После рецензирования: 17.02.2025
Принята к печати: 14.03.2025
В статье рассмотрены методические особенности учета биомассы и продуктивности на примере трех фоновых видов гипоарктических кустарников (Salix glauca, Betula exilis, Vaccinium uliginosum). Описан алгоритм проведения измерений и расчета фракционного состава надземной биомассы и годичной продуктивности с использованием показателя годичного прироста биомассы. Показано, что использование в качестве показателя продуктивности только верхушечного прироста побегов приводит к систематическому занижению показателя, поскольку не учитывает прироста древесины стволиков в процессе радиального прироста побегов. Между возрастом побегов и их длиной, а также диаметром основания побега не установлено значимой корреляции. В отношении массы побегов выявлено либо отсутствие связи, либо слабая связь с длиной побега и сильная связь с диаметром основания побега. Для всех видов характерно преобладание массы побегов, на долю которых приходится от 63 до 71% общей надземной фитомассы. Относительная скорость накопления фитомассы побегами в зависимости от вида колеблется от 7 до 10% в год. Установлена стабильность естественной влажности листовой фракции побегов исследуемых видов, среднее значение которой составило 61±2%.
Ключевые слова: полуостров Таймыр, биомасса, гипоарктические кустарники, возрастная структура, продуктивность, годичный прирост биомассы
Гипоарктические кустарники являются непременным компонентом растительности лесотундрового экотона (ЛТЭ), являясь как самостоятельным типом растительности, например, в форме зональных южных кустарниковых тундр (Александрова, 1977; Юрцев и др., 1978; Кожевников, 1988; Yurtsev, 1994; Поспелова, Поспелов, 2016; и др.), так и формируя соподчиненный ярус в северных редколесьях и рединах (Норин, 1978; Абаимов и др., 1991; Зырянова, Абаимов, 1997). В последнем случае их доля в общем балансе надземной биомассы может быть сопоставимой с древесной растительностью, а в рединах – в разы превышать ее (Игнатенко и др., 1973).
Кустарниковая растительность Арктики и Субарктики является одним из наиболее репрезентативных индикаторов последствий глобального потепления климата в этом регионе планеты, который выражается как в улучшении их роста и накопления фитомассы, так и продвижения кустарниковых тундр на север – эффект «позеленения» тундры (Forbes et al., 2010; Myers-Smith et al., 2011; Epstein et al., 2012; Frost, Epstein, 2014; Frost et al., 2014; Martin et al., 2017; Myers-Smith, Hik, 2018; Berner et al., 2020; Shevtsova et al., 2020; Seider et al., 2022; и др.).
Методы оценки продуктивности надземной фитомассы достаточно хорошо проработаны и широко используются в исследовании фитоценозов (Дылис и др., 1974; Whittaker, Marks, 1975; Андреева и др., 2002; и др.), в т.ч. тундровых (Александрова, 1958; Тихомиров и др., 1974; Цаценкин и др., 1978). Для оценки биомассы травяно-кустарничкового и мохово-лишайникового ярусов растительности широко используется «метод укосов», при котором на учетных площадках различного размера срезаются все растения, разделяются на отдельные компоненты, которые взвешивают, высушивают с последующим переводом в сухую фитомассу, которая в свою очередь служит основой для расчета содержания углерода, азота, других органогенных и минеральных элементов. В случае однолетних растений метод позволяет оценить как общую фитомассу, так и ее годичную продуктивность.
Для многолетних растений, к которым относятся кустарники, метод также применяется, в т.ч. и для тундровой растительности (Александрова, 1958; Игнатенко и др., 1973), но в этом случае вопрос о годичной продуктивности остается открытым. Для длительно живущих растений оценку годичной продуктивности проводят путем измерения годичных приростов верхушечных побегов (Базилевич и др., 1978; Scurlock et al., 2002). Однако изменение фитомассы происходит не только за счет прироста верхушечных побегов, но и, по аналогии с древесными растениями, посредством ее накопления в стволовой зоне в процессе радиального прироста стволиков побегов. Следовательно, чем крупнее растение, тем больше должна быть погрешность в определении годичной продуктивности при использовании традиционных подходов. Это предположение было принято в качестве тестируемой гипотезы в нашем исследовании.
Целью исследования являлось совершенствование существующих методических подходов для определения продуктивности кустарникового яруса как самостоятельного доминирующего структурного элемента в составе биогеоценоза (БГЦ), так и при его соподчиненном положении в составе БГЦ с доминированием древесного яруса. Помимо отработки методики в ходе исследования решались вспомогательные задачи, иллюстрирующие предлагаемый методический подход: а) определение возрастной структуры кустарниковой растительности; б) установление фракционного состава надземной фитомассы; в) расчет общей и годичной продуктивности; г) сравнение различных методов определения годичной продуктивности надземной фитомассы.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование проводилось на территории Таймыро-Среднесибирского биома южных гипоарктических тундр (Биоразнообразие биомов …, 2020) в охранной зоне участка «Ары-Мас» (Ары-Мас: Природные…, 1978) Таймырского государственного биосферного заповедника (рис. 1) в лиственничном редколесье из Larix gmelinii (Rupr.) Rupr. в районе кордона заповедника (72.46°с.ш., 101.94° в. д., ВУМ 24 м).\
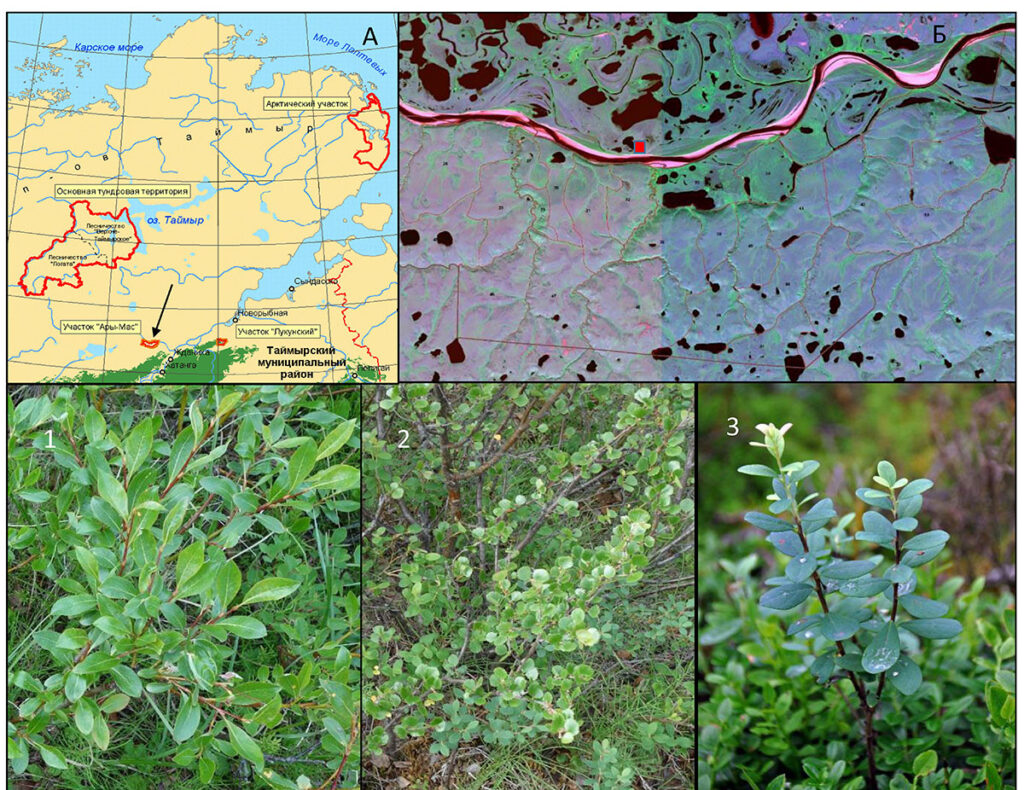
Рисунок 1. Местоположение и объекты исследования:
А – расположение участка «Ары-Мас» на территории полуострова Таймыр (красными контурами обозначены границы участков Таймырского ГПЗ), Б – местоположение отбора образцов на территории участка «Ары-Мас» (красный прямоугольник).
Исследуемые виды, формирующие кустарничковый ярус (здесь и далее по тексту):
1 – Salix glauca, 2 – Betula exilis, 3 – Vaccinium uliginosum)
В исследование были включены пять видов фоновых для данного региона гипоарктических кустарников: Betula exilis Sukaczev, Salix glauca L., Vaccinium uliginosum L., Salix lanata L., Duschekia fruticosa (Rupr.) Pouzar, для которых отбирались образцы побегов в пределах двух размерных групп: высотой до 0.5 м и более 0.5 м. Первая группа входит в состав травяно-кустарничкового яруса и представлена тремя видами: Betula exilis, Salix glauca, Vaccinium uliginosum. В ней отбирали по 10 образцов побегов каждого вида из среднего высотного диапазона яруса (20-40 см). Для каждого побега измеряли общую длину (высоту), диаметр у основания побега и диаметр на половине высоты. Побеги взвешивали непосредственно сразу после сбора в облиственном состоянии, после чего листья удаляли и определяли отдельно массу побега и листьев. Из удаленной листвы по каждому виду формировали навеску массой 10-15 г, которую в дальнейшем использовали для определения влажности листьев. Все собранные образцы упаковывали и высушивали до воздушно-сухого состояния и затем транспортировали для проведения измерений в лабораторных условиях.
Во вторую размерную группу вошли четыре вида: Betula exilis, Salix glauca, которые в данных условиях входят в состав, как травяно-кустарничкового яруса, так и кустарникового яруса, а также Salix lanata и Duschekia fruticosa. Все растения были представлены кустообразной формой, образованной несколькими побегами. Поскольку исследование носило прежде всего методический характер, то мы ограничились отбором одного среднего по размеру растения, для которого описывали/измеряли следующие характеристики: форма (кустообразная, древовидная), высота, проекция кроны в двух взаимно-перпендикулярных направлениях, количество отдельных побегов. Затем у каждого вида отбирали по два средних по размеру побега, для которых проводили весь комплекс измерений, аналогичный первой высотной группе.
В лабораторных условиях собранные образцы высушивали до абсолютно-сухого состояния и повторно измеряли те же морфометрические характеристики. Дополнительно отделяли и взвешивали прирост побегов за два последних года. По соотношению массы побегов (листьев) до и после их высушивания до абсолютно сухого состояния определяли влажность этих компонентов. Все дальнейшие расчеты проводили для абсолютно сухих образцов.
Для оценки годичной продуктивности надземной фитомассы побегов использовали показатель среднего годичного прироста побега, который определяли как отношение массы побега к его возрасту. Задача установления возраста оказалась весьма непростой в силу крайне незначительных размеров основания побегов, диаметр которых для первой возрастной группы составлял лишь несколько миллиметров.
Определение возраста проводили путем приготовления микроскопических препаратов из древесины побегов с использованием микротома санного типа. Перед резкой на микротоме образцы сухой древесины размягчались в дистиллированной воде. Затем с помощью микротома получали поперечные срезы древесины толщиной 10-15 мкм. Качество срезов оценивали под микроскопом и часть срезов отбраковывали. Наиболее качественные срезы использовали для приготовления временных микропрепаратов по известным методикам (Фурст, 1979; Бенькова, Швейнгрубер, 2004).
Для повышения контрастности изображения проводили окраску срезов 2%-ным раствором сафранина в течение 2-3 мин, при этом детали структуры становились более заметными. Окрашенные и промытые срезы помещали в каплю глицерина на предметное стекло. Окантовка препаратов была завершающей стадией подготовки качественных окрашенных срезов древесины, пригодных для микроскопического анализа.
Возраст побегов определяли путем подсчета количества годичных колец с использованием оборудования для компьютерного анализа изображений Axio Imager MAT, Version 04/14/2005 Karl Zeis Light AGMicroscopy с точностью 0.003 мм с применением специализированного программного обеспечения «Axio Vision» (Munro et al., 1996), которое позволяет определять линейные размеры и площади анатомических элементов в разных частях годичного кольца (рис. 2).
А |
Б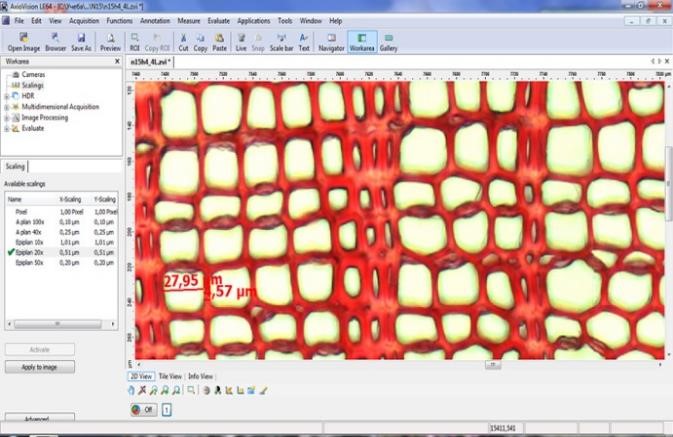 |
|
| В | 1.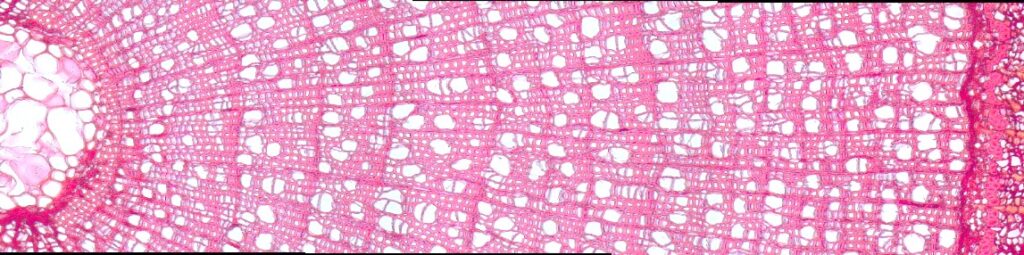 |
|
2.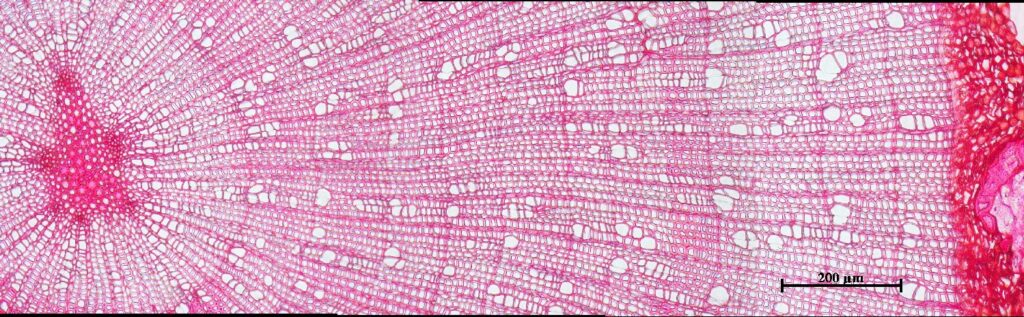 3. 3.
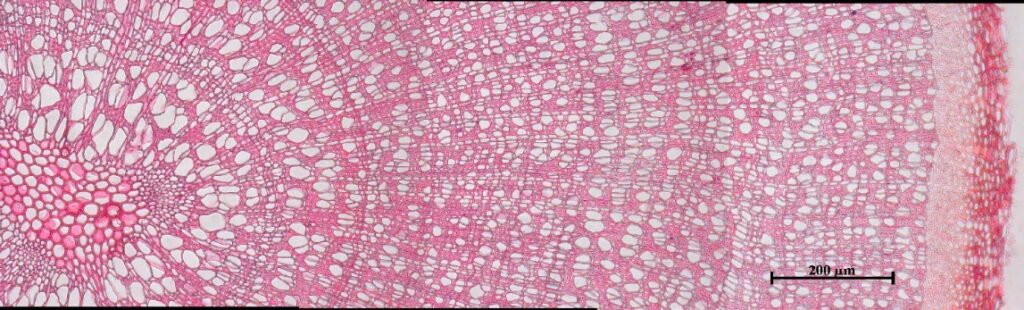 Рисунок 2. Определение возраста побегов:А — установка для компьютерного анализа изображений Axio Imager MAT, Karl Zeis Light AGMicroscopy; Б — интерфейс программного обеспечения «Axio Vision»; В — ширина и структура годичных колец побегов при 200-кратном увеличении (сверху-вниз: Salix glauca, Betula exilis, Vaccinium uliginosum) |
||
Для образцов первой размерной группы была выполнена статистическая обработка и регрессионный анализ соотношения морфометрических показателей с использованием программного продукта Statistica v.10.0.1011.0 (StatSoft …, 2011) и приложения Excel пакета офисных программ Microsoft Office. Для второй размерной группы, где количество отобранных образцов ограничивалось двумя побегами с каждого растения, приведены фактические результаты измерений, поскольку, на наш взгляд, они также представляют определенный интерес с точки зрения накопления данных по возрастной структуре и продуктивности кустарниковых видов в условиях Субарктики.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Характеристика морфометрические показателей. В процессе отбора образцов измеряли три показателя, характеризующих размеры побегов: длина (высота) побега, диаметр основания и диаметр на половине высоты. При этом предполагалось, что с использованием последнего показателя можно будет определить объем стволика с последующим расчетом биомассы с использованием формулы срединного сечения (формулы Губера (Анучин, 1982):
V = γ 1/2 × L (1)
где – V – объем стволика побега, γ1/2 — площадь поперечного сечения на середине длины и L – длина (высота) побега.
Первой проблемой, с которой пришлось столкнуться в процессе измерений, явилось определение срединного диаметра. Для видов с моноподиальным ветвлением (Betula exilis, Vaccinium uliginosum) это не составило большого труда, но в случае дихотомического типа ветвления (Salix glauca) задача оказалась затруднительной, поскольку у каждого ответвления был свой срединный диаметр. Кроме того, определение объема побегов с использованием стереометрических формул для такого типа ветвления теряет всякий смысл. В данном случае скорее подходит ксилометрический способ определения объема, что весьма затруднительно при полевых измерениях. Наконец, даже получив искомый объем с некоторым приближением, вопрос определения массы побега остается открытым, поскольку для большинства кустарниковых видов, в отличие от древесных, отсутствуют показатели плотности древесины, без которых переход от объема к массе невозможен. В результате в дальнейших исследованиях от измерения и использования диаметра на середине длины побега пришлось отказаться.
Для рядов распределения по диаметру характерна правосторонняя асимметрия, и лишь у Salix glauca отмечается смещение модального значения в левую часть распределения (рис. 3 А).
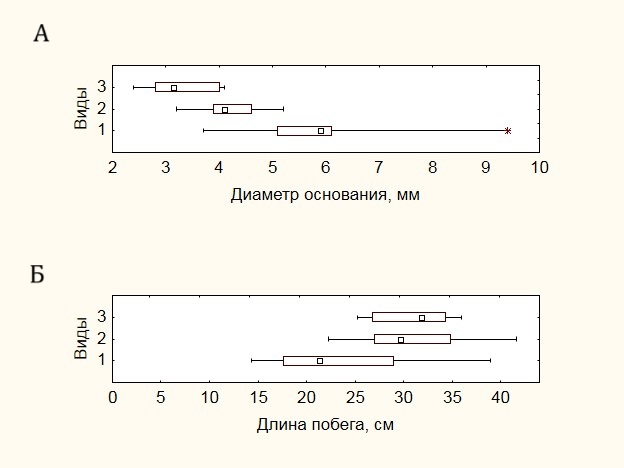
Рисунок 3. Характеристика рядов распределения морфометрических показателей:А — по диаметру основания побега, Б — по длине побега. Здесь и далее цифрами обозначены виды: 1 – Salix glauca, 2 – Betula exilis, 3 – Vaccinium uliginosum. Графическое построение выполнено по медианному значению
Для данного вида характерен и максимальный диапазон варьирования диаметров. Наибольший средний диаметр в основании побега – 6.0±1.5 мм отмечен для Salix glauca, наименьший – для Vaccinium uliginosum – 3.3±0.6 мм. Betula exilis в этом ряду занимает промежуточное положение – 4.2±0.7 мм. В отношении длины побегов наблюдается обратная картина (рис. 3 Б): наименьшее значение средней высоты отмечено у Salix glauca – 23.8±7.8 см, что связано с дихотомическим характером ветвления побегов данного вида, наибольшее у Vaccinium uliginosum – 30.9±4.0 см и практически такое же у Betula exilis — 30.8±5.6 см. Для Salix glauca и Betula exilis сохраняется аналогичный с диаметром основания характер распределения, в то время как для Vaccinium uliginosum распределение численностей по высоте характеризуется левосторонней асимметрией. В целом диапазон высот, в котором находится большинство особей исследуемых видов, варьирует от 15 до 40 см при среднем значении 28.5±6.7 см.
Несмотря на ограниченный размер выборки, изменчивость морфометрических показателей оказалась не слишком высокой. Для диаметров основания побега она варьирует от 16% у Betula exilis до 26% у Salix glauca, а в случае длины побегов — от 13% у Vaccinium uliginosum до 33% у Salix glauca.
Возрастная структура. По результатам подсчета годичных колец на микротомных срезах был определен возраст каждого побега и оценена возрастная структура отдельных видов (рис. 4). Наибольший средний возраст в рассматриваемом диапазоне высот отмечен у Salix glauca – 15±4 года, наименьший – у Vaccinium uliginosum – 11±4 года. Этому же виду присущ и наибольший интервал изменчивости возрастов, который составил 11 лет. Определенных тенденций в характере распределения по возрасту не отмечено. Для Vaccinium uliginosum характерна левосторонняя асимметрия, для Salix glauca – правосторонняя. В случае Betula exilis характер распределения максимально приближен к «нормальному», о чем свидетельствует совпадение среднего и медианного значений возраста. Несмотря на вариабельность возрастов, характерную в первую очередь для Vaccinium uliginosum и Salix glauca, средние значения возраста находятся в достаточно узком диапазоне – 13-15 лет, что свидетельствует о едином периоде формирования кустарничкового яруса. Наибольшая вариабельность возраста выявлена для Vaccinium uliginosum – 33%, наименьшая для Betula exilis – 17%.
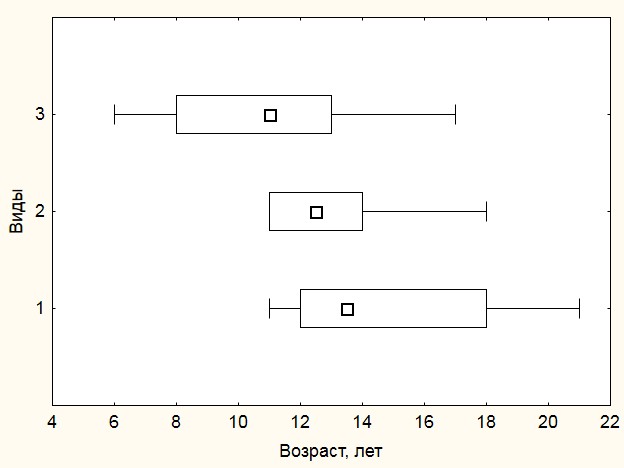
Рисунок 4. Характеристика возрастной структуры побегов
В ходе обработки данных была проанализирована теснота связи возраста побегов с диаметром основания (рис. 5 А) и их длиной (рис. 5 Б). В большинстве случаев отмечена положительная тенденция увеличения диаметра основания побега и его длины с увеличением возраста. Вместе с тем, для всех видов теснота связи оказалась незначимой (p ≫ 0.05), что не позволяет использовать морфометрические показатели для оценки возраста кустарников.
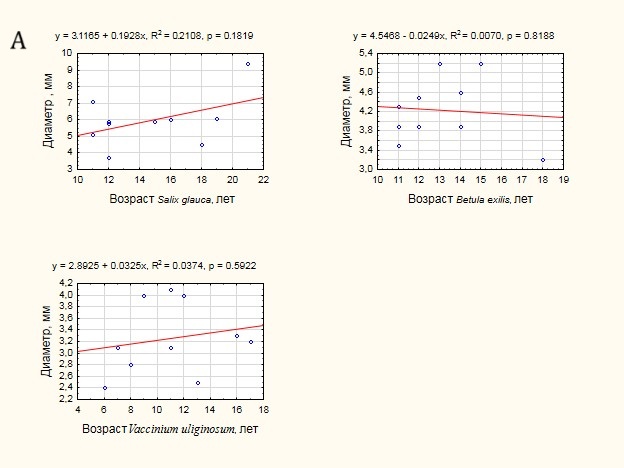
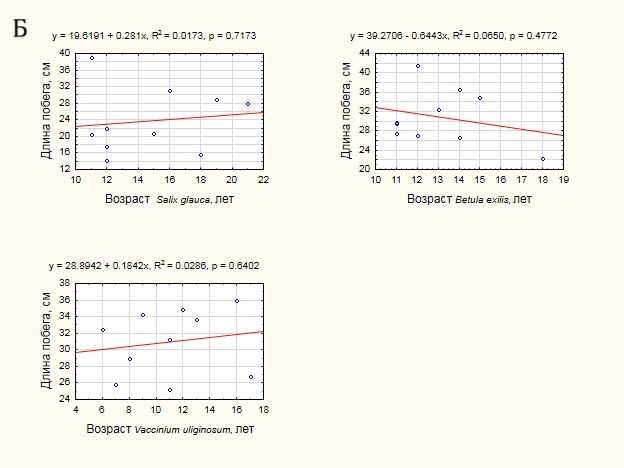
Рисунок 5. Оценка тесноты связи возраста побегов и морфометрических характеристик:А — с диаметром основания побега, Б — с длиной побега
Для образцов второй размерной группы приведены данные фактических измерений модельных побегов (табл. 1).
Таблица 1. Характеристика морфометрических показателей и возраста побегов размерной группы более 0.5 м
| № пп | Показатель | Ед. изм. | Вид | |||
| Salix lanata | Betula exilis | Salix glauca | Duschekia fruticosa | |||
| 1 | Высота растения | см | 130 | 110 | 78 | 210 |
| 2 | Площадь проекции кроны | м2 | 2.40 | 1.00 | — | 2.86 |
| 3 | Общее количество побегов | шт. | 17 | 9 | — | 14 |
| 4 | Количество модельных побегов | шт. | 2 | 2 | 2 | 2 |
| 5 | Длина побега | см | 129127-130 | 10293-110 | 6960-78 | 200190-210 |
| 6 | Диаметр основания побега | мм | 14.013.0-15.0 | 9.88.0-11.5 | 6.96.9-7.0 | 35.332.0-38.5 |
| 7 | Возраст, лет | лет | 12 | 1816-19 | 11 | 4342-44 |
Примечание. В числителе приведены средние значения, в знаменателе – их диапазон
Достаточно неожиданным результатом оказались близкие значения возраста побегов для обеих размерных групп, несмотря на значительные различия в морфометрических показателях, что свидетельствует о том, что один и тот же вид может достигать различных размеров в течение одинакового периода времени практически в одинаковых экологических условиях. Исключение составляет Duschekia fruticosa, отличающаяся несколько большими размерными показателями, но и значительно большим возрастом по сравнению с другими видами.
Структура надземной фитомассы и ее продуктивность. В результате обработки данных измерений для обеих размерных групп были получены значения общих и фракционных запасов надземной фитомассы для отдельных видов. В таблице 2 приведены данные для группы видов высотой до 0.5 м, а в таблице 3 – для видов, высота которых превышает 0.5 м.
Таблица 2. Характеристика фракционного состава надземной фитомассы побегов высотой до 0.5 м
| Показатель | Ед. изм. | Вид | ||
| Salix glauca | Betula exilis | Vaccinium uliginosum | ||
| Количество экземпляров | шт. | 10 | 10 | 10 |
| Фитомасса общая, в т.ч. | г | 5.8±3.8 | 2.3±1.0 | 1.6±0.7 |
| побеги | г | 3.7±2.6 | 1.7±0.8 | 1.2±0.6 |
| листья | г | 2.1±1.2 | 0.7±0.2 | 0.5±0.2 |
Примечание. Приведено среднее значение и среднеквадратическое отклонение,
Наибольшей массой в данной размерной группе во всех фракциях обладает Salix glauca, имеющая дихотомический тип ветвления побега, наименьшей – Vaccinium uliginosum. В отличие от морфометрических показателей фитомассе побегов присуща существенно большая изменчивость, которая колеблется от 49% у Betula exilis до 70% у Salix glauca.
Таблица 3. Характеристика морфометрических показателей и фракционного состава надземной фитомассы побегов высотой более 0.5 м
| Показатель | Ед. изм. | Вид | |||
| Salix lanata | Betula exilis | Salix glauca | Duschekia fruticosa | ||
| Количество экземпляров | шт. | 2 | 2 | 2 | 2 |
| Фитомасса общая, в т.ч. | г | 116.1
89.5-142.8 |
50.7
28.3-73.1 |
59.2
49.8-68.5 |
580.9
420.5-741.3 |
| Побеги | г | 84.5
67.4-101.6 |
40.3
25.3-55.3 |
24.3
19.4-29.0 |
444.1
365.8-522.4 |
| Листья | г | 31.6
22.1-41.2 |
10.4
3.0-17.8 |
34.9
30.4-39.5 |
136.8
54.7-218.9 |
Примечание. В числителе приведены средние значения, в знаменателе – их диапазон
По фитомассе у всех видов преобладает масса побегов, при этом соотношение доли побегов и листьев для видов, представленных в обеих размерных группах (Betula exilis и Salix glauca), принципиально не меняется. Вместе с тем доля массы побега для растений высотой менее 0.5 м оказалась несколько меньше по сравнению с группой, где высота растений превышает 0.5 м.
Для первой группы видов была проанализирована теснота связи между массой побега и морфометрическими характеристиками: длиной побега (рис. 6 А) и диаметром основания (рис. 6 Б). Достаточно неожиданным оказался результат соотношения массы побега и его длины. Связь оказалась незначимой (p> 0.05) в случае Betula exilis и Vaccinium uliginosum, что не позволяет использовать данный показатель для оценки массы побегов данных видов. Для Salix glauca связь – умеренная на 95% уровне значимости. Напротив, связь массы побега с диаметром его основания – сильная и значима для всех видов, несмотря на достаточно ограниченный размер выборки. В случае увеличения размера выборки можно ожидать получения адекватных моделей для расчета массы побегов по диаметру их основания.
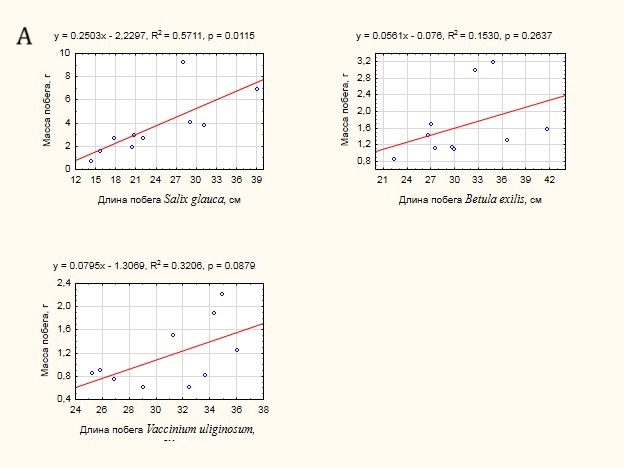
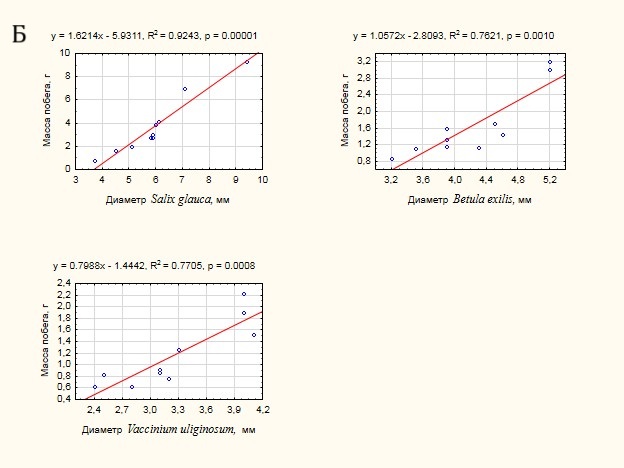
Рисунок 6. Оценка тесноты связи массы побега с морфометрическими характеристиками:
А – с длиной побега, Б – с диаметром основания
Имея массу побегов и их возраст, можно рассчитать среднюю скорость накопления фитомассы (средний годичный прирост):
ZM = Z / A (2)
где ZM – среднегодовой прирост массы побега, г год-1, M – масса побега, г, A — возраст побега, лет.
Разделив данный показатель на массу побега, получаем относительный среднегодовой прирост массы побега (PM) в процентах:
PM = (100× ZM ) / M (3)
Максимальное значение средней скорости накопления фитомассы побегов (среднегодовой прирост) без учета листьев выявлено для Salix glauca, 0.25±0.17 г год-1, что обусловлено кустистой формой побега данного вида (рис. 7).
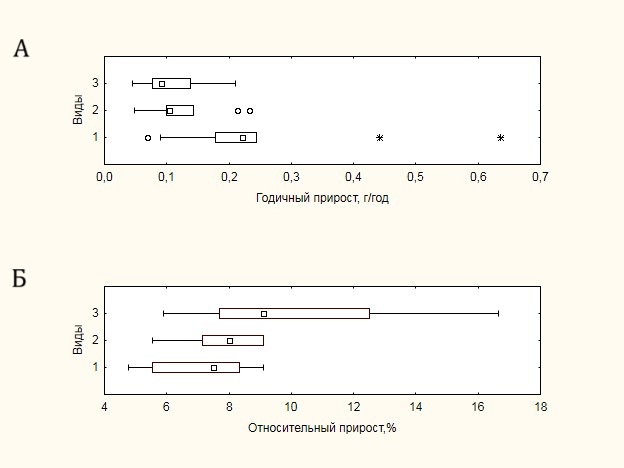
Рисунок 7. Годичный прирост побегов:
А – абсолютный, Б – относительный
Для Betula exilis и Vaccinium uliginosum, имеющих моноподиальный тип ветвления побега, значения среднего прироста побега оказались примерно в два раза ниже – 0.13±0.06 г год-1 и 0.11±0.05 г год-1 соответственно. Данному показателю для всех видов присуща достаточно высокая изменчивость: от 35% у Vaccinium uliginosum до 70% у Salix glauca.
В случае относительного прироста картина меняется: максимальный процент прироста отмечен у Vaccinium uliginosum — 10.1±3.5%, в то время как два других вида имеют близкие значения – 7.8±1.2% у Betula exilis и 7.2±1.7% у Salix glauca. При этом изменчивость относительного прироста уменьшается практически в два раза по сравнению с абсолютными значениями. Полученные значения относительной продуктивности примерно в два раза выше по сравнению с данными, полученными ранее для данного региона (BIODAT. Продуктивность экосистем…, 2006), что может быть связано как с методикой расчетов, так и с фиксируемым улучшением роста, обусловленным глобальным потеплением Арктики (Myers-Smith et al., 2011; Myers-Smith, Hik, 2018; Berner et al., 2020; Seider et al., 2022).
Очевидно, что показатели абсолютного прироста в значительной степени определяются размерами растения: чем оно крупнее, тем больше будет абсолютный прирост (табл. 4). Относительный прирост является более стабильным показателем и незначительно меняется для видов, представленных в обеих размерных группах. Несмотря на крупные размеры Duschekia fruticosa по сравнению с другими видами, показатель относительного годичного прироста побега оказался минимальным на фоне всех видов в обеих группах, что может быть обусловлено медленным ростом данного вида.
Таблица 4. Показатели годичной продуктивности побегов высотой более 0.5 м
| Показатель | Ед. изм. | Вид | |||
| Salix lanata | Betula exilis | Salix glauca | Duschekia fruticosa | ||
| Годичная продукция побегов | г/год | 7.0
5.6-8.5 |
2.4
1.3-3.5 |
2.2
1.8-2.6 |
10.4
8.3-12.4 |
| % | 8.3 | 5.8
5.3-6.3 |
9.1 | 2.0
2.3-2.4 |
|
Примечание. В числителе приведены средние значения, в знаменателе – их диапазон
Достаточно большой объем лабораторных исследований был связан с определением влажности образцов. В результате обработки данных по первой размерной группе было установлено, что влажность побегов специфична для различных видов (рис. 8).
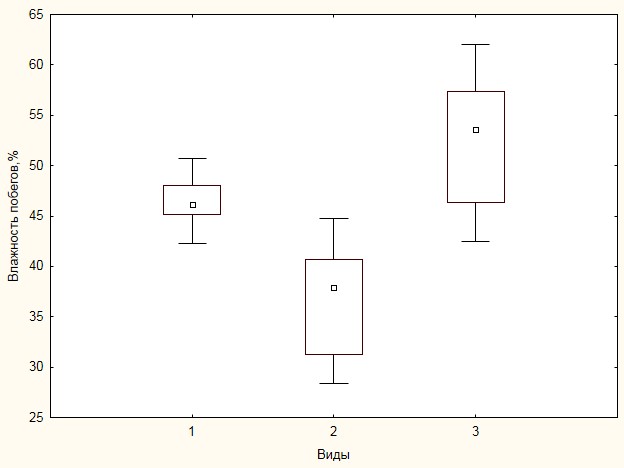
Рисунок 8. Характеристика влажности побегов высотой до 0.5 м
Наибольший процент влажности отмечен для Vaccinium uliginosum – 52.3±6.5%, далее в этом ряду следует Salix glauca – 46.5±6.5% и наименьшей влажностью обладают побеги Betula exilis – 37.0±5,4%. Для всех видов изменчивость влажности побегов не превышает 15% даже при столь малом объеме выборки. При ее увеличении можно ожидать более стабильного значения показателя, который может быть использован в расчетах в качестве константы, что в свою очередь позволит существенно уменьшить количество образцов, необходимых для обработки в лабораторных условиях в пользу их увеличения при производстве измерений в полевых условиях. При увеличении размеров растений принципиальных различий влажности побегов не выявлено (табл. 5).
Таблица 5. Влажность побегов высотой более 0.5 м
| Показатель | Ед. изм. | Вид | |||
| Salix lanata | Betula exilis | Salix glauca | Duschekia fruticosa | ||
| Влажность побегов в коре | % | 47.1
46.0-48.2 |
41.6
41.0-42.3 |
50.7
48.7-52.8 |
46.0
45.0-47.5 |
| Влажность побегов без коры | % | 42.5
40.8-44.2 |
38.1
37.1-39.0 |
41.3
35.3-47.4 |
— |
Примечание. В числителе приведены средние значения, в знаменателе – их диапазон
Влажность фракции листьев определялась в целом для вида путем получения массы навески из нескольких образцов. Принципиальных различий как между видами, так и между размерными группами не выявлено (рис. 9), среднее значение составило 61.4±2.3%. В случае Duschekia fruticosa можно предположить, что с увеличением размера растения влажность листовой фракции несколько увеличивается. Но этот вывод и приведенные данные требуют более тщательной проверки, поскольку получены по результатам измерения одной навески по каждому виду и здесь мы их приводим скорее в качестве предварительной оценочной информации.
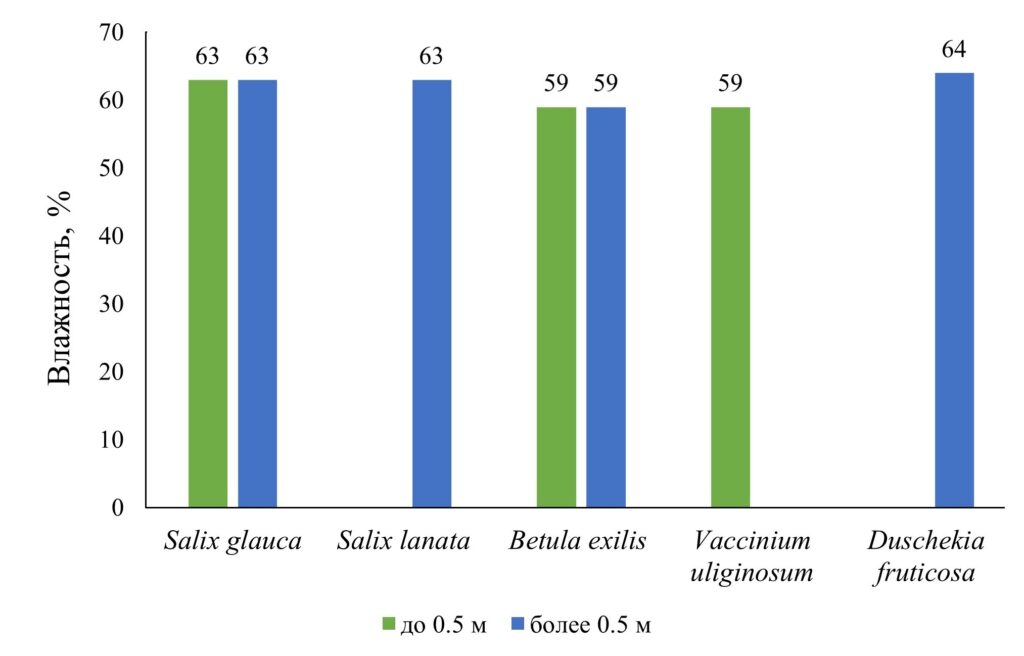
Рисунок 9. Влажность фракции листьев по размерным группам
Традиционным способом определения годичного прироста фитомассы для кустарниковых видов является измерение массы побегов текущего года (Андреева и др., 2002). В рамках исследовании мы провели сравнительный анализ годичной продукции надземной биомассы побегов, полученной обоими способами: по средней массе приростов побегов и по среднему годичному приросту для группы видов высотой до 0.5 м (рис. 10).
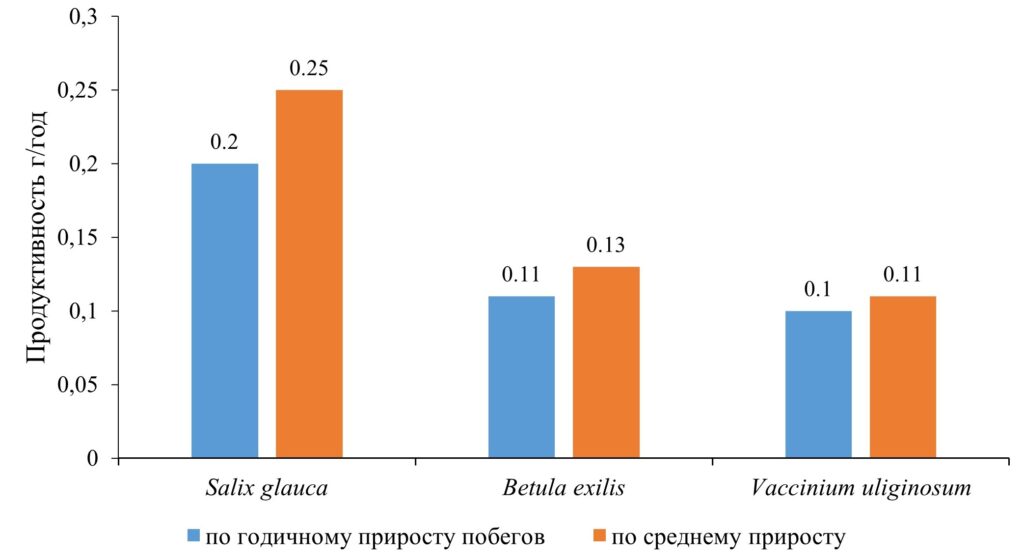
Рисунок 10. Сравнение различных способов определения годичной продуктивности побегов
Для всех видов отмечено превышение годичной продукции, рассчитанной на основании среднего прироста побега, по сравнению с рассчитанной по средней массе приростов побегов. Причем различие возрастает по мере увеличения массы растения и, например, в случае Salix glauca достигает 25%. По мере увеличения размера растений можно предположить дальнейшее увеличение этого различия.
Все вышеперечисленные результаты касались только одиночных растений, вернее их побегов, но поскольку обычно стоит задача оценки продуктивности на единице площади, то требуется переход от одиночного побега к растению в случае, если его образуют несколько побегов, затем к учетной площадке и от нее уже к единице площади.
В случае кустарников, представленных несколькими побегами, алгоритм измерений, включает выбор нескольких средних по размеру побегов (2-3), их измерение, затем пересчет полученных результатов на 1 растение. Далее подсчитывается количество растений на единице учетной площади, вычисляются фитомасса и продуктивность на учетной площади и пересчитываются на единицу площади, чаще на 1 м2 (Berner et al., 2024). Если побеги, образующие растение, сильно различаются по высоте, то более точные данные можно получить, предварительно разделив растение на высотные ярусы, из которых затем уже отбирать модельные образцы.
Проведенное исследование подтвердило тестируемую гипотезу о занижении оценки биомассы побегов кустарниковых видов при использовании для расчетов только годичного прироста побегов в силу того, что данный метод не учитывает радиального прироста по диаметру самого побега. С учетом полученного результата более точным способом учета годичной продукции биомассы кустарниковых видов растений является учет их общей массы и определение возраста растений. Вполне вероятным представляется получение поправочных коэффициентов к результатам, рассчитанным по годичному приросту побегов, в зависимости от вида и массы всего растения по мере накопления данных о среднем приросте.
Кроме того, определенный оптимизм внушает стабильность относительного прироста фитомассы отдельных видов, который по мере накопления данных может быть использован для оценки годичной продукции по массе растения, которая по результатам исследований имеет тесную связь с диаметром основания побега, что позволяет существенно сократить объем натурных измерений и использовать для расчетов регрессионные уравнения.
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования протестирован алгоритм оценки фитомассы и годичной продуктивности трех фоновых видов гипоарктических листопадных кустарников (Betula exilis, Salix glauca, Vaccinium uliginosum), формирующих травяно-кустарничковый ярус в лиственничном редколесье на северной границе распространения древесной растительности. Показано, что традиционный метод учета продуктивности фитомассы по годичному приросту побегов приводит к систематическому занижению годичной продукции за счет недооценки доли продукции, накапливаемой в побегах в процессе прироста по диаметру. Для более точной оценки скорости накопления биомассы предлагается использовать показатель среднегодового прироста, определяемого как отношение общей фитомассы побега (растения) к его возрасту.
Результаты проведенного анализа не выявили достоверной связи между возрастом побега и его размерными характеристиками: диаметром основания и длиной побега, что не позволяет использовать эти параметры для оценки возраста. Вместе с тем выявлена тесная связь массы побегов с диаметром основания, что дает возможность использовать для оценки фитомассы надземной части побега регрессионные уравнения. В то же время установлено наличие слабой связи или ее отсутствие на 95% уровне значимости массы побегов с высотой растения.
Дополнительно в рамках проведенного исследования были получены предварительные оценки общей надземной биомассы побегов рассматриваемых видов и ее фракционной структуры, скорости накопления биомассы, проведен сравнительный анализ различных методов оценки годичной продуктивности надземной биомассы.
Результаты выполненного исследования имеют важное методическое значение, поскольку использованный подход позволяет более корректно оценивать годичную продукцию кустарничковых видов, исходя из их количества на единице площади и среднегодовой скорости накопления фитомассы отдельными растениями. Полученные значения фракционного состава фитомассы модальных побегов позволяют с приемлемой точностью вычислять фитомассу всего растения, а путем подсчета их количества на учетной площади — общую фитомассу кустарникового яруса и его годовую продуктивность.
Полученные результаты по мере накопления дополнительных данных будут использованы для уточнения конверсионных коэффициентов фракционного состава надземной фитомассы кустарниковой растительности, а также построения регрессионных моделей.
БЛАГОДАРНОСТИ
Работа выполнена в рамках реализации важнейшего инновационного проекта государственного значения «Разработка системы наземного и дистанционного мониторинга пулов углерода и потоков парниковых газов на территории Российской Федерации, обеспечение создания системы учета данных о потоках климатически активных веществ и бюджете углерода в лесах и других наземных экологических системах» (рег. № 123030300031-6).
Авторы выражают признательность Объединенной дирекции заповедников Таймыра (ФГБУ «Заповедники Таймыра»), а также сотрудникам государственного природного биосферного заповедника «Таймырский» за поддержку и содействие в проведении исследований.
КОНФЛИКТ ИНТЕРЕСОВ
Авторы сообщают об отсутствии конфликта интересов.
ИСТОЧНИК ДАННЫХ
Результаты первичных измерений побегов размерной группы до 0.5 м, использованные при подготовке статьи приведены в Приложении 1, а размерной группы более 0.5 м – в табл.1.
СПИСОК ЛИТЕРАТУРЫ
Абаимов А. П., Бондарев А. И., Коротков И. А., Софронов М. А. Эколого- географические особенности притундровых лесов Сибири // Эколого-географические проблемы сохранения и восстановления лесов Севера. Тез. докл. Всесоюзн. научн. конференции. Архангельск, 1991. С. 67–69.
Александрова В. Д. Опыт определения надземной и подземной массы растительности в Арктической тундре // Ботанический журнал. 1958. Т. XIII. № 12. С. 1748–1761.
Александрова В. Д. Геоботаническое районирование Арктики и Антарктики. Л.: Наука, 1977. 187 с.
Андреева Е. Н., Баккал И. Ю., Горшков В. В., Лянгузова И. В., Мазная Е. А., Нешатаев В. Ю., Ставрова Н. И., Ярмишко В. Т., Ярмишко М. А. Методы изучения лесных сообществ. СПБ: НИИХимии СПбГУ, 2002. 240 с.
Анучин Н. П. Лесная таксация. Изд. 5-е, дополн. М: Лесная промышленность, 1982. 552 с.
Ары-Мас: Природные условия, флора и растительность самого северного в мире лесного массива / Под ред. Б. Н. Норина. Л.: Наука. Ленингр. отделение, 1978. 192 с.
Базилевич Н. И., Титлянова А. А., Смирнов В. В., Родин Л. Е., Нечаева Н. Т., Левин Ф. И. Методы изучения биологического круговорота в различных природных зонах. М.: Мысль, 1978. 185 с.
Бенькова В. Е., Швейнгрубер Ф.Х. Анатомия древесины растений России (Атлас для идентификации древесины деревьев, кустарников, кустарничков, полукустарников и деревянистых лиан России). Бирменсдорф: Хаупт, 2004. 456 с.
БИОДАТ. Продуктивность экосистем Северной Евразии. 2006. URL: https://www.biodat.ru/db/prod/index.htm (дата обращения 05.11.24).
Биоразнообразие биомов России. Равнинные биомы / Под ред. Г. Н. Огуреевой. М.: ФГБУ «ИГКЭ», 2020. 623 с.
Дылис Н. В., Карпов В. Г., Цельникер Ю. Л. Изучение высшей растительности как компонента биогеоценоза // Программа и методика биогеоценологических исследований / Н. В. Дылис (отв. ред.). М.: Наука, 1974. С. 73–80.
Зырянова О. А., Абаимов А. П. Типологическая структура растительного покрова // Леса Красноярского Заполярья. Новосибирск: Наука, 1997. С. 55–95.
Игнатенко И. В., Кнорре А. В., Ловелиус Н. В., Норин Б. Н. Запасы фитомассы в типичных растительных сообществах лесного массива «Ары-Мас» // Экология. 1973. № 3. С. 36–43.
Кожевников Ю. П. О южных тундрах // Ботанический журнал. 1988. Т. 73. № 1. С. 65–74.
Норин Б. Н. Растительный покров центральной части урочища // Ары-Мас: Природные условия, флора и растительность самого северного в мире лесного массива. Л.: Наука, 1978. С. 133–162.
Поспелова Е. Б., Поспелов И. Н. Флора подзоны южных тундр полуострова Таймыр // Растительный мир Азиатской России. 2016. № 1 (21). С. 80–88.
Тихомиров Б. А., Шамурин В. Ф., Норин Б. Н. Изучение тундровых биогеоценозов // Программа и методика биогеоценологических исследований / Н. В. Дылис (отв. ред.). М.: Наука, 1974. С. 257–260.
Фурст Г. Г. Методы анатомо-гистохимического исследования растительных тканей. М.: Наука, 1979. 156 с.
Цаценкин И. А., Савченко И. В., Дмитриева С. И. Методические указания по экологической оценке кормовых угодий тундровой и лесной зон Сибири и Дальнего Востока по растительному покрову. М.: ВНИИ кормов, 1978. 301 с.
Юрцев Б. А., Толмачев А. И., Ребристая О. В. Флористическое разграничение и разделение Арктики // Арктическая флористическая область. Л.: Наука, 1978. С. 9–104.
Berner L. T., Massey R., Jantz P., Forbes B. C., Macias-Fauria M., …, & Goetz S. J. Summer warming explains widespread but not uniform greening in the Arctic tundra biome // Nature Communications. 2020. Vol. 11. No. 1. Article 1. P. 1–12. DOI: 10.1038/s41467-020-18479-5
Berner L. T., Orndahl K. M., Rose M., Tamstorf M., Arndal M. F., …, & Goetz S. J. The arctic plant aboveground biomass synthesis dataset // Scientific Data. 2024. Vol. 11. Article 305. P. 1–13. DOI: 10.1038/s41597-024-03139-w
Epstein H. E., Raynolds M. K., Walker D. A., Bhatt U. S., Tucker C. J., Pinzon J. E. Dynamics of aboveground phytomass of the circumpolar Arctic tundra during the past three decades // Environmental Research Letters. 2012. Vol. 7. No. 1. Article 015506. P. 1–12. DOI: 10.1088/1748-9326/7/1/015506
Forbes B. C., Fauria M. M., Zetterberg P. Russian Arctic warming and ‘greening’ are closely tracked by tundra shrub willows // Global Change Biology. 2010. Vol. 16. P. 1542–1554. DOI: 10.1111/j.1365-2486.2009.02047.x
Frost G. V., Epstein H. E. Tall shrub and tree expansion in Siberian tundra ecotones since the 1960s // Global Change Biology. 2014. Vol. 20. P. 1264–1277. DOI: 10.1111/gcb.12406
Frost G. V., Epstein H. E., Walker D. A. Regional and landscape-scale variability of Landsat-observed vegetation dynamics in northwest Siberian tundra // Environmental Research Letters. 2014. Vol. 9. Article 025004. P. 1–11. DOI: 10.1088/1748-9326/9/2/025004
Martin A. C., Jeffers E. S., Petrokofsky G., Myers-Smith I., Macias-Fauria M. Shrub growth and expansion in the Arctic tundra: an assessment of controlling factors using an evidence-based approach // Environmental Research Letters. 2017. Vol. 12. Article 085007. P. 1–13. DOI: 10.1088/1748-9326/aa7989
Munro M. A. R., Brown P. M., Hughes M. K., Garcia E. M. R. Image analysis of tracheid dimensions for dendrochronological use // Radiocarbon. 1996. P. 843–853.
Myers-Smith I. H., Forbes B. C., Wilmking M., Hallinger M., Lantz T., …, & Hik D. S. Shrub expansion in tundra ecosystems: dynamics, impacts and research priorities // Environmental Research Letters. 2011. Vol. 6 (4). Article 045509. DOI: 10.1088/1748-9326/6/4/045509
Myers-Smith I. H., Hik D. S. Climate warming as a driver of tundra shrubline advance // Journal of Ecology. 2018. Vol. 106 (2). P. 547–560. DOI: 10.1111/1365-2745.12817
Scurlock J. M. O., Johnson K., Olson R. J. Estimating net primary productivity from grassland biomass dynamics measurements // Global Change Biology. 2002. Vol. 8. P. 736–753.
Seider J. H., Lantz T. C., Hermosilla T., Wulder M. A., Wang J. A. Biophysical determinants of shifting tundra vegetation productivity in the Beaufort Delta region of Canada // Ecosystems 2022. Vol. 25 (7). P. 1435–1454. DOI: 10.1007/s10021-021- 00725-6
Shevtsova I., Heim B., Kruse S., Schröder J., Troeva E. I., Pestryakova L. A., Zakharov E. S., Herzschuh U. Strong shrub expansion in tundra-taiga, tree infilling in taiga and stable tundra in central Chukotka (north-eastern Siberia) between 2000 and 2017 // Environmental Research Letters. 2020. Vol. 15. Article 085006. P. 1–21. DOI: 10.1088/1748-9326/ab9059
StatSoft, Inc. 2011. STATISTICA (data analysis software system), version 10. Available on https://statistica.software.informer.com/10.0/ (дата обращения 10.09.24).
Whittaker R. H., Marks P. L. Methods of Assessing Terrestrial Productivity // Primary Productivity of the Biosphere / H. Lieth, R. H. Whittaker (Eds.). Ecological Studies. Vol. 14. Springer, Berlin, Heidelberg, 1975. P. 55–118. DOI: 10.1007/978-3-642-80913-2_4
Yurtsev B. A. Floristic division of the Arctic // Journal of Vegetation Science. 1994. Vol. 5. P. 765–776. DOI: 10.2307/3236191
Приложение 1.
Результаты первичных измерений побегов в размерной группе до 0.5 м
| № побега | Длина побега, см | Диаметр основания побега. мм | Возраст, лет | Масса при естественной влажности, г | Абсолютно сухая масса, г | ||||
| побег с листьями | побега без листьев | листья | побег | прирост предпоследнего года | прирост последнего года | ||||
| Salix glauca | |||||||||
| 1 | 20.6 | 5.9 | 15 | 7.9 | 5.4 | 2.5 | 3.0 | 0.12 | 0.14 |
| 2 | 29.0 | 6.1 | 19 | 10 | 7.1 | 2.9 | 4.1 | 0.12 | 0.12 |
| 3 | 22.0 | 5.9 | 12 | 7.5 | 5.0 | 2.5 | 2.7 | 0.18 | 0.22 |
| 4 | 39.0 | 7.1 | 11 | 19.9 | 13.7 | 6.2 | 7.0 | 0.30 | 0.30 |
| 5 | 31.0 | 6.0 | 16 | 9.8 | 7.5 | 2.3 | 3.9 | 0.18 | 0.16 |
| 6 | 28.0 | 9.4 | 21 | 24.5 | 17.4 | 7.1 | 9.3 | 0.50 | 0.62 |
| 7 | 17.6 | 5.8 | 12 | 8.8 | 5.6 | 3.2 | 2.8 | 0.12 | 0.28 |
| 8 | 20.4 | 5.1 | 11 | 6.4 | 3.6 | 2.8 | 2.0 | 0.12 | 0.18 |
| 9 | 15.6 | 4.5 | 18 | 5.1 | 3.1 | 2.0 | 1.6 | 0.10 | 0.10 |
| 10 | 14.3 | 3.7 | 12 | 2.3 | 1.5 | 0.8 | 0.8 | 0.04 | 0.02 |
| Навеска листьев | 15.3 | 5.6 | |||||||
| Betula exilis | |||||||||
| 1 | 22.3 | 3.2 | 18 | 1.9 | 1.2 | 0.7 | 0.9 | 0.07 | 0.11 |
| 2 | 27.5 | 4.3 | 11 | 2.3 | 1.6 | 0.7 | 1.1 | 0.05 | 0.12 |
| 3 | 29.6 | 3.9 | 11 | 3.3 | 2.1 | 1.2 | 1.2 | 0.04 | 0.11 |
| 4 | 32.5 | 5.2 | 13 | 6.2 | 4.9 | 1.3 | 3.0 | 0.02 | 0.08 |
| 5 | 36.6 | 3.9 | 14 | 3.3 | 2.2 | 1.1 | 1.3 | 0.02 | 0.14 |
| 6 | 27.0 | 4.5 | 12 | 3.9 | 2.9 | 1.0 | 1.7 | 0.07 | 0.22 |
| 7 | 29.8 | 3.5 | 11 | 2.8 | 1.6 | 1.2 | 1.1 | 0.18 | 0.27 |
| 8 | 26.6 | 4.6 | 14 | 3.2 | 2.3 | 0.9 | 1.4 | 0.03 | 0.17 |
| 9 | 41.6 | 3.9 | 12 | 4.1 | 2.7 | 1.4 | 1.6 | 0.05 | 0.16 |
| 10 | 34.8 | 5.2 | 15 | 7.2 | 5.1 | 2.1 | 3.2 | 0.07 | 0.13 |
| Навеска листьев | 9.7 | 4.0 | |||||||
| Vaccinium uliginosum | |||||||||
| 1 | 25.8 | 3.1 | 7 | 2.2 | 1.6 | 0.6 | 0.9 | 0.02 | 0.04 |
| 2 | 32.4 | 2.4 | 6 | 1.6 | 1.4 | 0.2 | 0.6 | 0.06 | 0.04 |
| 3 | 34.9 | 4.0 | 12 | 5.9 | 4.5 | 1.4 | 2.2 | 0.14 | 0.16 |
| 4 | 26.8 | 3.2 | 17 | 2.0 | 1.3 | 0.7 | 0.8 | 0.08 | 0.08 |
| 5 | 33.6 | 2.5 | 13 | 2.1 | 1.5 | 0.6 | 0.8 | 0.08 | 0.04 |
| 6 | 29.0 | 2.8 | 8 | 1.7 | 1.0 | 0.7 | 0.6 | 0.04 | 0.04 |
| 7 | 25.2 | 3.1 | 11 | 2.1 | 1.5 | 0.6 | 0.9 | 0.08 | 0.04 |
| 8 | 36.0 | 3.3 | 16 | 3.6 | 2.3 | 1.3 | 1.3 | 0.10 | 0.06 |
| 9 | 31.2 | 4.1 | 11 | 3.9 | 2.9 | 1.0 | 1.5 | 0.10 | 0.06 |
| 10 | 34.3 | 4.0 | 9 | 5.1 | 4.1 | 1.0 | 1.9 | 0.14 | 0.10 |
| Навеска листьев | 6.6 | 2.7 | |||||||
Рецензент: к.б.н. Геникова Н. В.





