- DOI: 10.31509/2658-607x-202582-168
- УДК 581.526.524
Ценопопуляционный анализ и прогнозы развития сосняков- зеленомошников в Неруссо-Деснянском полесье
![]()
© 2025 Н. В. Короткова1, О. И. Евстигнеев1,2
1 Центр по проблемам экологии и продуктивности лесов им. А. С. Исаева РАН
Россия, 117997 Москва, ул. Профсоюзная, 84/32, стр. 14
2Государственный природный биосферный заповедник «Брянский лес»
Россия, 242180, Брянская обл., Суземский р-н, ст. Нерусса, ул. Заповедная, 2
E-mail: natalya-1998-494@mail.ru
Поступила в редакцию: 15.05.2025
После рецензирования: 06.06.2025
Принята к печати: 10.06.2025
На основе ценопопуляционного анализа синузии деревьев проведена оценка сукцессионного состояния сосняков-зеленомошников и даны прогнозы их развития в Неруссо-Деснянском Полесье. Проанализирован онтогенетический состав ценопопуляций разных видов деревьев в этих сообществах. Показано, что сосняки-зеленомошники, которые расположены в непосредственной близости от генеративных деревьев ели обыкновенной, постепенно преобразуются в монодоминантные разновозрастные ельники-зеленомошники. Установлено, что сосняки-зеленомошники, которые расположены вдали от источников семян ели и вблизи от плодоносящего дуба черешчатого, способны сначала преобразоваться в хвойно-широколиственных леса с большим участием дуба. Однако по мере выпадения из сообщества сосны и дуба, которые не способны возобновляться в лесах теневого типа, формируются олигодоминантные елово-широколиственные леса с преобладанием теневыносливых видов деревьев – ели обыкновенной, клена остролистного и липы сердцевидной.
Ключевые слова: ценопопуляции деревьев, сукцессионное состояние сообществ, ельники-зеленомошники, хвойно-широколиственные леса, Pinus sylvestris, Picea abies, Quercus robur
В настоящее время в растительном покрове полесий господствуют монодоминантные сосняки (Гроздов, 1951; Юркевич и др., 1977; Растительность …, 1980; Морозова, 1999; Панченко, 2013). Эти сообщества – пирогенный вариант деградации полидоминантных елово-широколиственных лесов. Сосняки сформировались под воздействием частых пожаров, которые непреднамеренно или умышленно провоцировал человек (Кулешова и др., 1996; Евстигнеев и др., 1999; Смирнова, Коротков, 2001; Восточноевропейские …, 2004б; Евстигнеев, 2009; Evstigneev, Korotkov, 2016б; Smirnova et al., 2017). Пожары на территории полесий полностью «съедают» подстилку и перегнойно-аккумулятивный горизонт, оставшийся песчаный субстрат становится недоступным для поселения многих видов растений. Пожары в сочетании с рубками уничтожили большую часть видового состава синузии деревьев. Сохранились преимущественно сосна обыкновенная, ель обыкновенная, березы бородавчатая и пушистая, дуб черешчатый (Евстигнеев, 2009, 2010). Лесоводы относят эти виды к олиготрофам (Ремезов, Погребняк, 1965; Погребняк, 1968). У перечисленных деревьев, кроме ели, также выражена огнестойкость, которая определяется особенностями их биологии и экологии: 1) толстой корой стволов, которая защищает камбий от высоких температур (дуб, сосна, береза); 2) формированием порослевых побегов из спящих почек на ксилоподии после утраты ствола подростом (дуб, береза); 3) поверхностно-стержневой и поверхностно-стержнево-якорной корневыми системами, у которых часть корней расположена ниже прожигаемого субстрата (дуб, сосна, береза); 4) высоко поднятой кроной у взрослых деревьев (сосна); 5) способностью семян быстро и обильно появляться в послепожарных сообществах или анемохорным (сосна, береза), или зоохорным способами (дуб); 6) приживанием проростков на оголенном песчаном субстрате (сосна). Широкому распространению сосняков на песчаных почвах, начиная с первых циркуляров Лесного департамента XIX века, немало способствовали лесокультурные мероприятия, направленные на выращивание деловой и быстрорастущей древесины сосны, которая хорошо растет на бедных почвах (Цветков, 1957).
Опыт предшествующих исследователей показал, что направления сукцессионных преобразований сосняков зависят от взаимоотношений видов деревьев и характера семенного потока с окружающих территорий. Лесоводы считают, что в основе сукцессионного переформирования подроста и древостоя сосновых лесов лежат взаимоотношения преимущественно трех видов: сосны, дуба и ели (Сукачев, 1938; Мелехов, 1944; Морозов, 1949; Ткаченко, 1952; Атрохин, 1982 и др.). Одновременно эти авторы отмечают, что на ход развития сообществ могут оказывать влияние и другие виды деревьев (например, клен остролистный и липа сердцевидная). Исход борьбы сосны, дуба, ели и других видов зависит прежде всего от их теневыносливости и скорости роста по высоте. Виды выстраиваются в следующий ряд по увеличению теневыносливости: сосна, дуб, ель, липа, клен (Evstigneev, 2018; Евстигнеев, 2022). Во всех возрастах наибольшая скорость роста по высоте характерна для сосны (Загреев и др., 1992). Это позволяет сосне на пожарищах, вырубках, заброшенных пашнях и в культурах обогнать другие внедряющиеся виды и быстро сформировать древостой. Кроме того, исследователи полагают, что на взаимоотношения видов деревьев и, соответственно, на сукцессионные изменения сообществ существенное влияние оказывает характер семенного потока с окружающих территорий (Мелехов, 1944; Коротков, 1992; Восточноевропейские …, 1994, 2004б; Евстигнеев, 2010; Евстигнеев, Воеводин, 2012).
Широкие возможности для оценки сукцессионного состояния лесных сообществ и прогнозов их развития дает ценопопуляционный анализ. Этот подход разработан в отечественной демографии растений (Смирнова и др., 1987, 1989, 1991; Восточноевропейские …, 1994, 2004а; Браславская, 2019). В связи с этим в работе поставлена задача – дать прогнозы развития сосняков-зеленомошников на основе ценопуляционного анализа синузии деревьев.
РАЙОН, ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили в Неруссо-Деснянском Полесье. Эта территория по ботанико-географическому районированию относится к зоне широколиственных лесов, которая принадлежит Полесской подпровинции Восточно-европейской провинции Европейской широколиственно-лесной области (Растительность …, 1980). В качестве объекта исследования выбрали ценозы одного типа леса – сосняка-зеленомошника (Pinetum pleuroziosum) по классификации И. Д. Юркевича и др. (1977). Эти сообщества преобладают в Неруссо-Деснянском Полесье, они занимают более половины территории (Евстигнеев, 2010). Подстилающая порода сосняка-зеленомошника – песок; глубина грунтовых вод – около полутора метров; местообитание по эдафической сетке П. С. Погребняка (1968) – влажная суборь, B3. В ярусе трав преобладают Calamagrostis epigeios (L.) Roth, Convallaria majalis L., Dryopteris carthusiana (Vill.) H. P. Fuchs, Maianthemum bifolium (L.) F. W. Schmidt, Melampyrum pratense L., Molinia caerulea (L.) Moench, Pteridium aquilinum (L.) Kuhn, Solidago virgaurea L., Trientalis europaea L., Vaccinium myrtillus L., V. vitis—idaea L. В напочвенном покрове доминируют зеленые мхи – Pleurozium schreberi (Brid.) Mitt., Dicranum scoparium Hedw., Polytrichum juniperinum Hedw.
Для ценопопуляционного анализа заложили восемь временных пробных площадей размером от 0.5 до 1.5 га. Все деревья по высоте поделили на две фракции: первая – до 50 см; вторая – более 50 см. Численности особей первой фракции учитывали на площадках по 100 м2 в десятикратной повторности, второй – сплошным пересчетом на всей пробе. Полученные данные пересчитывали на 1 га. Для каждой особи указывали биологический возраст, ее онтогенетическое (возрастное) состояние: j – ювенильное; im – имматурное; v – виргинильное; g1, g2, g3 – молодое, средневозрастное (зрелое) и старое генеративное; s – сенильное. При этом опирались на ранее опубликованные работы по биологическому возрасту ели обыкновенной (Романовский, 2001), сосны обыкновенной (Евстигнеев, 2014), дуба черешчатого (Евстигнеев, Короткова, 2024), липы сердцевидной (Чистякова, 1979) и других видов деревьев (Evstigneev, Korotkov, 2016а).
На основе полученных данных построили онтогенетические спектры ценопопуляций деревьев. Онтогенетический спектр – это распределение особей в ценопопуляции по биологическому возрасту. На спектрах онтогенетические состояния выражены в абсолютных числах и представлены в виде гистограмм. Полученные гистограммы сравнивали с характерным онтогенетическим спектром. У всех видов деревьев характерный спектр – поночленный и левосторонний. В нем представлены особи всех онтогенетических состояний, а по численности преобладают растения прегенеративного периода. Ценопопуляции с таким спектром способны к самоподдержанию, поскольку отличаются устойчивым оборотом поколений. Полночленный и левосторонний спектр, как правило, свойственен климаксным и субклимаксным сообществам. Его структура определяется биологическими свойствами вида (Смирнова и др., 1987; Восточноевропейские …, 1994). Ценопопуляции, испытавшие на себе катастрофические воздействия (рубки, пожары и т.п.), называют сукцессионными. Их онтогенетические спектры в разной степени отклонены от характерного (Заугольнова и др., 1988). Все разнообразие сукцессионных спектров объединяются в три типа: 1) инвазионные, которые представлены лишь прегенеративными (иногда и молодыми генеративными) растениями; 2) регрессивные, состоящие из постгенеративных растений (иногда в спектре присутствуют g2 и g3 особи); 3) фрагментарные, представленные единичным числом особей одного или двух онтогенетических состояний. Между этими типами спектров существует множество переходов (Восточноевропейские …, 2004а).
В большинстве сообществ, где проводили исследования, были видны следы от пожаров – обугленные стволы деревьев и огневые подсушины в их основании, а также угли в верхних горизонтах почвы. Датировку низовых пожаров определяли по абсолютному возрасту подроста Picea abies и побегов формирования (поросли) лиственных деревьев. При оценке возраста древесных растений опирались на методы, разработанные А. А. Корчагиным (1960). После низовых пожаров подрост ели полностью погибает. Это связано с его слабой огнестойкостью (Мелехов, 1944). Однако в районе исследования ель после пожаров начинает быстро восстанавливаться, поскольку ее отдельные генеративные особи сохраняются по влажным окраинам многочисленных болот. Возраст подроста ели определяли по числу мутовок на стволе. Давность последних низовых пожаров выявляли также по возрасту побегов формирования у подроста деревьев и кустарников – Sorbus aucuparia, Malus sylvestris, Quercus robur и др. Побеги формирования у этих видов сразу начинают развиваться после пожаров из спящих почек, расположенных на ксилоподии, который сохраняется в нижней части поврежденного растения. Возраст побегов формирования определяли по смене годичных приростов. В каждом из ценозов провели от 18 до 40 датировок последних низовых пожаров.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Наибольшее значение для преобразования сосняков имеет дальность расположения источников семян ели, дуба и других широколиственных деревьев. По условиям инвазий сосняки-зеленомошники Неруссо-Деснянского Полесья делятся на две группы (рис. 1). Первая – сообщества, которые расположены в непосредственной близости от источников семян ели, дуба, и находящиеся вдали от генеративных широколиственных деревьев (более 2000 м).
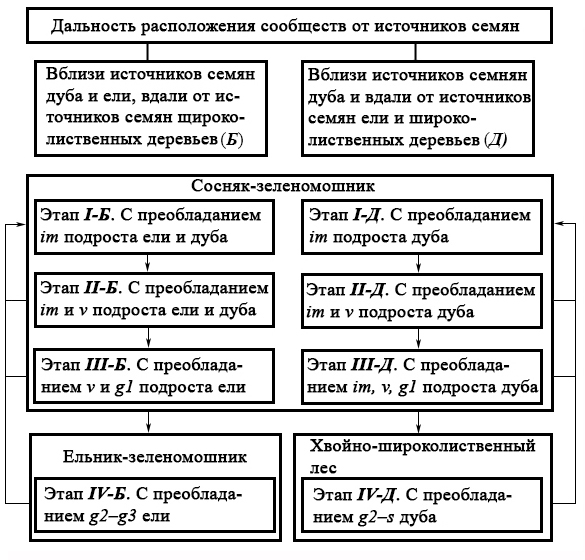
Рисунок 1. Направления развития сосняков-зеленомошников на разном удалении от источников семян ели, дуба и других широколиственных деревьев
Обозначения: стрелки вниз – направления развития сообществ, определяемые популяциями деревьев; стрелки вверх – преобразования сообществ при низовых пожарах; римские цифры – этапы сукцессионных преобразований;
онтогенетическое (возрастное) состояние: j – ювенильное; im – имматурное; v – виргинильное; g1, g2, g3 – молодое, средневозрастное (зрелое) и старое генеративное; s – сенильное
Эта группа в работе обозначена буквой «Б». Вторая группа – сосняки, расположенные в непосредственной близости от плодоносящих дубов и далее 300 м от источников семян ели и других широколиственных деревьев. Эта группа в работе обозначена буквой «Д» (рис. 1). Несмотря на слабую огнестойкость, ель широко представлена в Неруссо-Деснянском Полесье. Это определяется следующими обстоятельствами. Во-первых, генеративные деревья влаголюбивой ели в большом числе сохранились в местообитаниях с пониженной горимостью: по окраинам сырых западин и на низинных болотах. Во-вторых, при воспроизводстве лесов на песчаных почвах активно создаются еловые культуры (Евстигнеев, 2009), которые отличаются относительно малой требовательностью к богатству почвы (Ремезов, Погребняк, 1965) и быстрым ростом после десяти лет (Ткаченко, 1952). Источники семян клена и липы – хвойно-широколиственные леса. Они сохранились в основном по бортам и поймам ручьев, малых и средних рек (Евстигнеев, 2004; 2010).
Ценопопуляционный анализ и прогнозы развития сосняков-зеленомошников, расположенных в непосредственной близости от источников семян ели и дуба
В сукцессионных преобразованиях сосняков, которые расположены в непосредственной близости от генеративных деревьев дуба и ели, выделяются четыре этапа по степени развития внедряющегося подроста этих видов: I) сосняк-зеленомошник с преобладанием im подроста ели и дуба; II) сосняк-зеленомошник с доминированием im и v подроста ели и дуба; III) сосняк-зеленомошник с господством v–g1 елей; IV) ельник-зеленомошник с преобладанием g2–g3 елей (рис. 1).
Этап I–Б. Сосняк-зеленомошник с преобладанием im подроста ели и дуба (рис. 2). Ярус древостоя – культуры сосны, возраст которых составляет 85 лет. В сообществе последний низовой пожар был 14 лет назад. Это влияет на онтогенетический (возрастной) состав ценопопуляций деревьев.
У сосны прерывисто неполночленный онтогенетический спектр с высокой численностью особей. Взрослая фракция представлены g2-особями нормальной жизненности и g1-растениями низкой жизненности. В молодой фракции в большом числе присутствует j и im подрост (рис. 3, I). Большая численность подроста сосны обусловлена тем, что этот вид хорошо возобновляется на минеральном субстрате, который остается после низового пожара (Санников, Санникова, 1985). К древостою в небольшом числе примешиваются березы бородавчатая и пушистая. Береза бородавчатая по характеру распределения онтогенетических состояний ближе к регрессивному типу спектра, в котором в большом числе представлены g3 и s особи. Подрост березы отсутствует из-за его слабой теневыносливости (Евстигнеев, 2022). У более теневыносливой березы пушистой онтогенетический спектр – прерывисто неполночленный с низкой численностью особей. Представлены только единичные g3 деревья, а по нарушениям напочвенного покрова встречаются редкие j особи.

Рисунок 2. Сосняк-зеленомошник с преобладанием im подроста ели и дуба (этап I–Б)
Обозначения: А – общий вид сообщества, в ярусе подроста представлены дуб черешчатый и ель обыкновенная, на заднем фоне видны генеративные ели; Б – имматурная ель обыкновенная второй подгруппы (im2) рядом с обгоревшим стволом сосны обыкновенной во время низового пожара 2011 г.; В, Г – имматурный дуб первой подгруппы (im1) в квазисенильном состоянии (синоним – «торчок»), в основании подроста заметна верхняя часть ксилоподия; Д – имматурный дуб первой подгруппы (im1) семенного происхождения. Даты фотографий: А, Б – начало марта 2025 г., В, Г, Д – август 2024 г.
Наиболее активно в сообщество внедряются только два вида деревьев – дуб черешчатый и ель обыкновенная. Это связано с тем, что генеративные особи дуба и ели в большом числе сохранились в окружающих сообществах. Известно, что желуди дуба заносят сойки, а семена ели – ветер (Евстигнеев и др., 2013, 2017). Кроме того, относительно высокая освещенность под сосновым пологом позволяет подросту дуба и ели беспрепятственно приживаться и развиваться. За 14 лет, прошедших после последнего низового пожара, у дуба сформировалась ценопопуляция инвазионного типа с высокой численностью j или im особей (рис. 3, I). Относительно высокая плотность дуба определяется не только деятельностью сойки, но и тем, что одна четверть подроста пережила низовой пожар 2011 года. Она представлена квазисенильными особями (рис. 2, В, Г). У этих растений, испытавших огневое поражение, погибла надземная часть, однако из спящих почек на ксилоподии появились новые порослевые побеги.
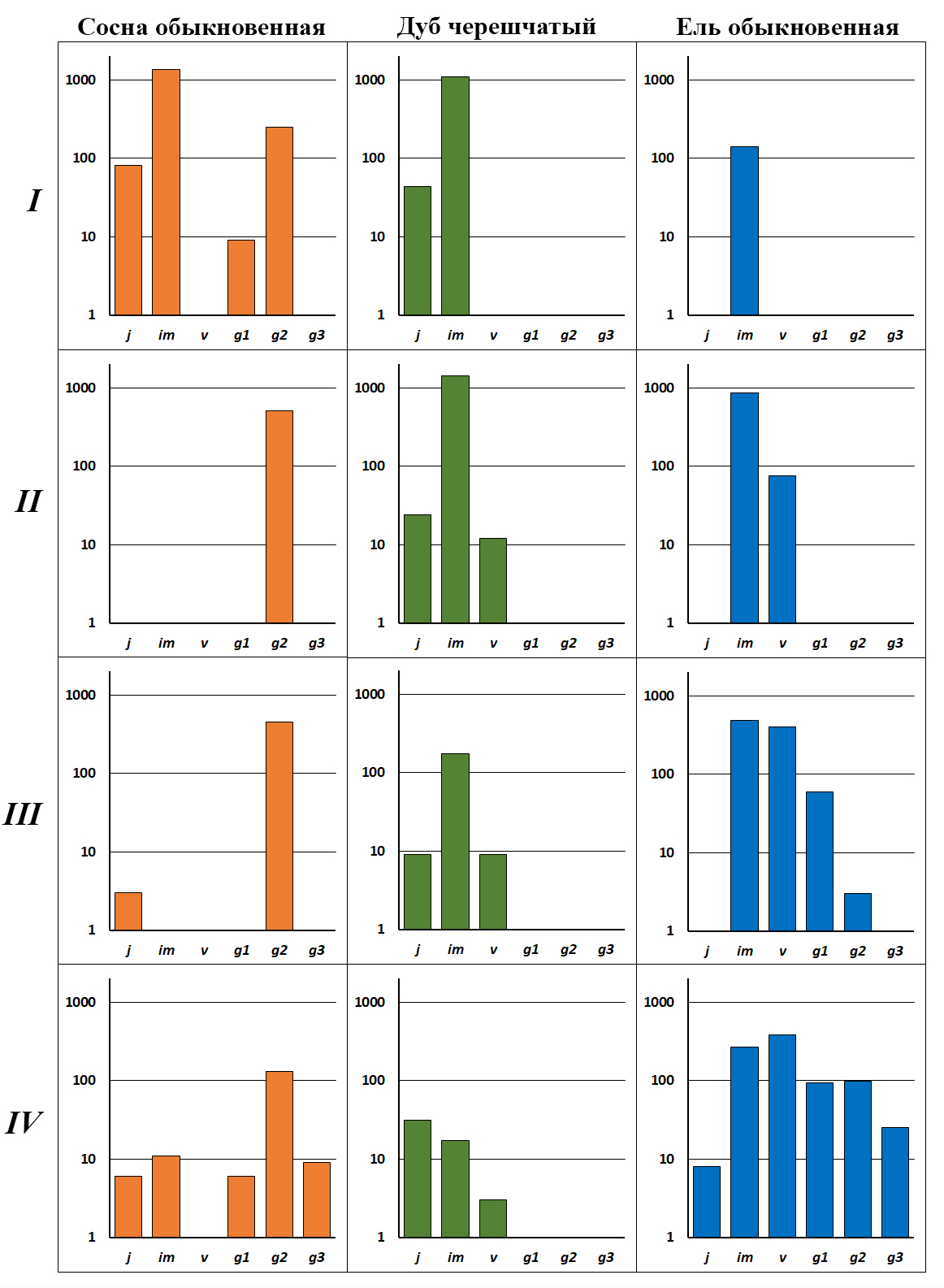
Рисунок 3. Онтогенетический состав ценопопуляций деревьев в сосняках-зеленомошниках, рядом с которыми расположены генеративные деревья дуба черешчатого и ели обыкновенной
Обозначения. Римские цифры – этапы формирования ценопопуляций (см. текст). По оси х – онтогенетические состояния, по оси y – число особей на 1 га в логарифмическом масштабе
У ели также сформировалась ценопопуляция инвазионного типа, в которой представлены im особи (рис. 3, I). Однако ее численность почти на порядок меньше, чем у дуба. Это связано с тем, что весь подрост ели выгорел после низового пожара 2011 года. Известно, что ель отличается слабой пожароустойчивостью: тонкая кора не защищает камбий от высоких температур, корневая система поверхностного типа полностью погибает во время пожара, крона со смолистой хвоей, которая опущена до земли, быстро уничтожается огнем. Кроме того, вновь внедряющийся подрост ели предъявляет большую требовательность к влажности субстрата, чем дуб. Это связано с тем, что ель развивает только поверхностную корневую систему, а дуб – поверхностно-стержневую. Кроме того, лесоводы показали, что сосна и дуб в пятилетнем возрасте испаряют за сутки 75% воды, которая содержится в их тканях, а ель – 150–200% (Погребняк, 1968). Другими словами, ель в течение суток должна дважды восполнить свой запас воды. В подросте также отмечены осина и клен остролистный, однако их популяции фрагментарного типа, они представлены единичными j или im особями. Их судьбу в сообществе предсказать невозможно. Ценопопуляционный анализ показывает, что будущее сосняка-зеленомошника зависит от взаимоотношения двух видов в подросте – дуба черешчатого и ели обыкновенной.
Этап II-Б. Сосняк-зеленомошник с преобладанием im и v подроста ели и дуба (рис. 4). Ярус древостоя представлен культурами сосны, возраст которых составляет 75 лет. В сообществе последний низовой пожар был 40 лет назад. Это отражается на возрастной структуре ценопопуляций деревьев.
Популяция сосны представлена только g2-особями нормальной жизненности, подрост полностью отсутствуют (рис. 3, II). Последнее связано с тем, что в сообществе, по сравнению с предыдущим этапом, хорошо сформирован ярус кустарников и подроста деревьев: его сомкнутость возросла с 0.4 до 0.7, а высота – с одного метра до пяти. Этот ярус затеняет и вытесняет светолюбивый подрост сосны. К древостою в небольшом числе примешивается g3 береза бородавчатая, ее малочисленный подрост представлен im и v особями крайне низкой жизненности.
Ярус подроста формируют главным образом два вида деревьев – дуб черешчатый и ель обыкновенная (рис. 4). За 40 лет отсутствия пожаров, а также благодаря активному заносу семян с окружающих территорий, численность инвазионных ценопопуляций этих видов существенно возросла: у дуба – на 300 особей, а ели – в семь раз. В подросте этих деревьев сформировались v-особи (рис. 3, II). Значительное увеличение численности влаголюбивой ели связано с тем, что почвенный покров в сообществе, видимо, стал меньше иссушаться вследствие появления развитого яруса подроста. Ценопопуляции других деревьев (березы пушистой, груши обыкновенной, клена остролистного и яблони лесной) относятся к фрагментарному типу. Они представлены одним–двумя онтогенетическими состояниями с небольшой численностью особей. Ценопопуляционный анализ показал, что на этом этапе развития сообщества у сосны полностью прекращается формирование молодого поколения, а участие дуба и ели в ярусе подроста возрастает.

Рисунок 4. Сосняк-зеленомошник с преобладанием im и v подроста ели и дуба (этап II–Б)
Обозначения: А, В – общий вид сообщества, в ярусе подроста представлены дуб черешчатый и ель обыкновенная; Б – виргинильная ель первой подгруппы (v1); Г – виргинильный дуб первой подгруппы (v1). Даты фотографий: А, Б – начало марта 2025 г.; В – октябрь 2024 г., Г – июнь 2024 г.
Этап III–Б. Сосняк-зеленомошник с v—g1 елью и единичным подростом дуба (рис. 5). В древостое, как и в предыдущем сообществе, господствует сосна, которая выросла из культур. Возраст сосны – 100 лет. В сообществе последний низовой пожар был 70 лет назад. Это серьезно повлияло на структуру внедряющего подроста.
Популяция сосны представлена в основном g2-особями нормальной жизненности. По сравнению с предыдущим этапом часть генеративных деревьев вывалилась. В популяции сосны иногда появляется единичный подрост, однако из-за низкой освещенности его жизнь ограничена j состоянием (рис. 3, III). Календарный возраст этих особей не бывает больше двух–трех лет. К древостою в небольшом числе примешиваются березы бородавчатая и пушистая. У светолюбивой березы бородавчатой онтогенетический спектр – регрессивный, а у более теневыносливой березы пушистой – регрессивно-инвазионный. Это свидетельствует о том, что первое поколение этих берез, появившееся сто лет назад, практически полностью вывалилось из сообщества. У теневыносливой березы пушистой в небольшом числе прижилось второе поколение, однако оно представлено единичным im и v подростом.

Рисунок 5. Сосняк-зеленомошник с v-g1 елью и с единичным подростом дуба (этап III–Б)
Обозначения: А – общий вид сообщества, в ярусе подроста представлена преимущественно ель обыкновенная; Б – ствол молодой генеративной ели (g1); В – на переднем плане – виргинильный дуб первой подгруппы (v1) низкой жизненности. Дата фотографий – начало марта 2025 г.
На этом этапе развития сообщества в ярусе подроста господствует только один вид – теневыносливая ель обыкновенная (рис. 5). Численность ее популяции осталась на прежнем уровне – 941 особь на га. Однако структура онтогенетического спектра существенно поменялась: из инвазионной преобразовалась в нормальную левостороннюю. В ней отсутствуют только j и g3-особи (рис. 3, III). Для формирования g3 деревьев недостаточно 70 лет, которые прошли после последнего низового пожара. Отсутствие j-особей можно объяснить тем, что в лесах молодое поколение ели предпочитает приживаться на валежинах (Мелехов, 1989; Кикеева и др., 2022). Однако в рассматриваемом ценозе валеж еще не появился, поскольку сообщество находится на первых этапах восстановления. В связи с этим семена ели прорастают только периодически – во влажные годы c частыми дождями.
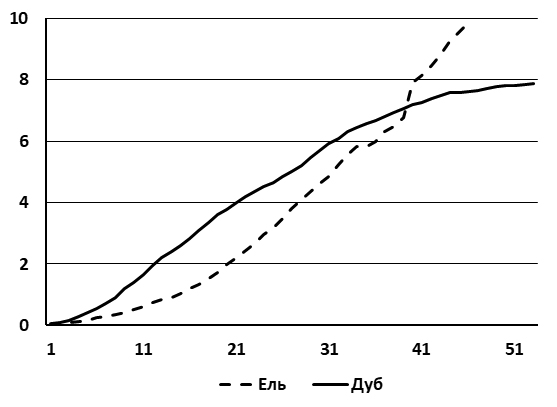
Рисунок 6. Ход роста по высоте виргинильных особей дуба черешчатого и ели обыкновенной под пологом сосняка-зеленомошника. По горизонтальной оси – возраст в годах, по вертикальной – высота деревьев в метрах. Кривые получены на основе анализа 15 особей каждого вида
В этом сообществе популяция дуба начинает деградировать. Это определяется особенностями роста дуба и ели по высоте. В сосновых лесах приросты по высоте у дуба на первых этапах онтогенеза превышают еловые. Однако после 40 лет дуб начинает уступать по высоте ели (рис. 6). В результате над подростом дуба формируется затеняющий ярус из v и g1 елей. Развитие инвазионной ценопопуляции светолюбивого дуба останавливается, число его особей сокращается почти в восемь раз (рис. 3, III), а жизненность сохранившегося подроста становится низкой и крайне низкой. Фрагментарные ценопопуляции груши, клена и яблони, которые встречались на предыдущих этапах, в этом ценозе полностью исчезли из-за недостатка света.
Этап IV–Б. Ельник-зеленомошник с g2–g3 елью и единичным подростом дуба (рис. 7). В ярусе древостоя начинает преобладать ель. Это свидетельствует о том, что в ходе сукцессионных преобразований меняется тип леса: сосняк-зеленомошник преобразуется в ельник-зелемномошник. Сосна – это культуры, которые были посажаны 95 лет назад. За это время в сообществе не отмечены низовые пожары. В древостое в небольшом числе присутствует береза бородавчатая.

Рисунок 7. Ельник-зеленомошник с g2–g3 елью и единичным подростом дуба (этап IV–Б)
Обозначения: А – вид сообщества, в ярусах древостоя и подроста представлена преимущественно ель обыкновенная; Б – окно на месте вывала g3 сосны (на заднем плане) и основание ствола от сломленной g3 ели (на переднем плане); В – основание ствола g3 ели и вывал ели; Г – основание ствола от сломленного g1 дуба низкой жизненности (справа) и вывал ели (вдали). Дата фотографий – начало марта 2025 г.
У сосны и березы бородавчатой прерывисто неполночленный онтогенетический спектр, в котором полностью отсутствуют v особи. По сравнению с предыдущим этапом в ценопопуляции сосны появились g3 особи (рис. 3, IV). Однако число генеративных особей этого вида уменьшилась в три раза. Эта связано с выходом ели в верхний ярус, она затеняет крону светолюбивой сосны и вытесняет ее из сообщества. К этому времени у ели сформировалась полночленная левосторонняя ценопопуляция, в которой представлены особи всех онтогенетических состояний (рис. 3, IV), что свидетельствует об устойчивом обороте поколений. Этому способствует появление валежа, на котором начали приживаться молодые особи ели. Упавшие и перегнивающие стволы деревьев – это благоприятный субстрат для приживания проростков. Исследователи пишут, что валеж отличается повышенной аэрацией и влажностью, меньшими колебаниями температуры, большей доступностью элементов минерального питания и отсутствием корневой конкуренции со стороны взрослых деревьев (Кикеева и др., 2022). Перегнивающие стволы осваивает подрост и других видов деревьев (березы, сосны, осины).
Темный полог, который на этом этапе развития сообщества сформирован из генеративных елей, определяет дальнейшую деградацию ценопопуляций дуба. Численность особей его инвазионной ценопопуляции, в которой представлены j, im и v особи (рис. 3, IV), уменьшается в четыре раза по сравнению с предыдущим сообществом. Весь подрост дуба относится к низкой и крайне низкой жизненности. Сойки продолжают заносить в сообщество желуди. Они прорастают, однако редко доживают до im1 состояния из-за дефицита света.
Анализ развития ценопопуляций дуба черешчатого в сосняках-зеленомошниках, рядом с которыми расположены генеративные деревья ели обыкновенной, выявил несколько моментов. Дуб сразу после низовых пожаров начинает активно внедряться в сообщества и в течение первых 40–50 лет формирует популяции инвазионного типа с высокой численностью особей. Однако после 40–50 лет интенсивно вселяющаяся теневыносливая ель обгоняет светолюбивый дуб по высоте, затеняет его и постепенно вытесняет из сообщества. По мере старения и выпадения из ценоза генеративной сосны на месте сосняков-зеленомошников постепенно образуются монодоминантные ельники-зеленомошники. В этих сообществах у ели формируются ценопопуляции с устойчивым оборотом поколений. Такие сообщества с ограниченным видовым составом деревьев и с нормальным оборотом поколений в их ценопопуляциях называют диаспорическим субклимаксом (Смирнова, Торопова, 2008).
Ценопопуляционный анализ и прогнозы развития сосняков-зеленомошников, расположенных рядом с плодоносящими дубами и вдали от источников семян ели, клена и липы
В этой части работы рассматриваются сосняки-зеленомошники, которые расположены в непосредственной близости от плодоносящих деревьев дуба и на расстоянии 300 и более метров от генеративных елей и широколиственных деревьев (клена, липы и др.). В сукцессионных преобразованиях этих сообществах выделяются четыре этапа по степени развития внедряющегося дуба: I) сосняк-зеленомошник с преобладанием im подроста дуба; II) сосняк-зеленомошник с господством im и v подроста дуба; III) сосняк-зеленомошник с доминированием im–g1 особей дуба; IV) хвойно-широколиственный лес с превосходством g2–s дубов (рис. 1).
Этап I–Д. Сосняк-зеленомошник с преобладанием im подроста дуба (рис. 8). Ярус древостоя представлен культурами сосны, возраст которых составляет 75 лет. В сообществе последний низовой пожар был 16 лет назад в 2009 году. Это отражается на онтогенетической (возрастной) структуре ценопопуляций деревьев. Во взрослой фракции сосны господствуют g2 деревья нормальной жизненности и в меньшем числе представлены g1 особи низкой жизненности, которые отстали в своем развитии. В подросте сосны присутствует большое число j и im особей (рис. 9, I). Все они прижились в сообществе на минеральном субстрате, который появился после низового пожара 2009 года. Однако из-за слабой теневыносливости весь подрост сосны относится к низкой жизненности, который развивается не дальше im онтогенетического состояния (рис. 8, Б). В древостое также присутствует береза бородавчатая, у которой ценопопуляция регрессивно-инвазионного типа с малым числом особей.

Рисунок 8. Сосняк-зеленомошник с преобладанием im подроста дуба (этап I–Д)
Обозначения: А – общий вид сообщества, в ярусе подроста преобладает дуб, встречается единичная ель; Б – в подросте обычна имматурная сосна первой подгруппы (im1); В – обгоревший ствол сосны во время низового пожара 2009 г. Дата фотографий – октябрь 2024 г.
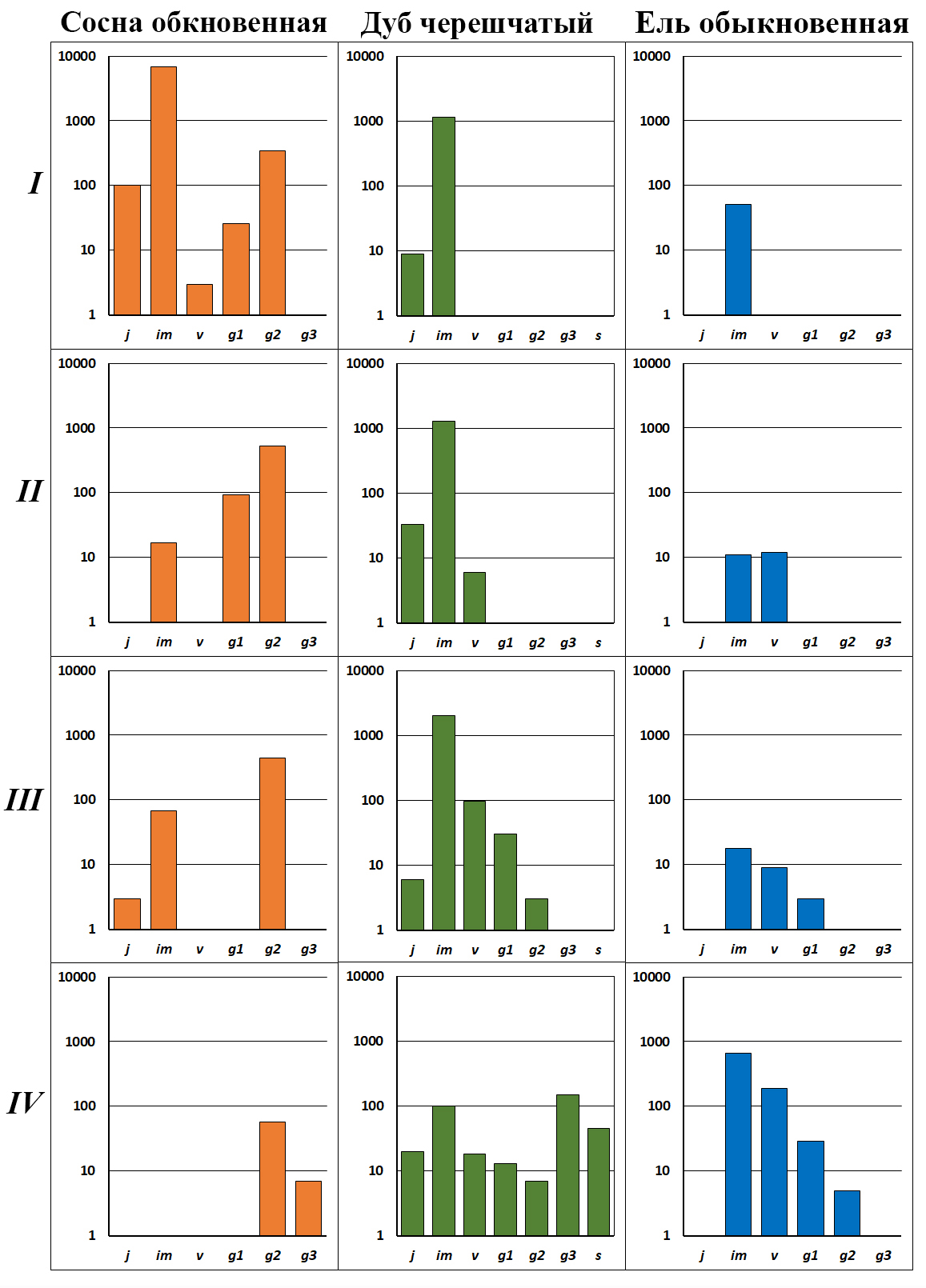
Рисунок 9. Онтогенетический состав ценопопуляций деревьев в сосняках-зеленомошниках, которые расположены вблизи плодоносящих дубов черешчатых и вдали от генеративных елей обыкновенных и других широколиственных деревьев
Обозначения. Римские цифры – этапы формирования ценопопуляций (см. текст). По оси х – онтогенетические состояния, по оси y – число особей на 1 га в логарифмическом масштабе
Дуб к моменту изучения сформировал ценопопуляцию инвазионного типа с высокой численностью im особей (рис. 9, I). Половина этого подроста пережила низовой пожар и в настоящее время представлена квазисенильными растениями. В сообществе присутствует ель. Однако ее ценопопуляция фрагментарного типа, в которой представлены единичные im особи. Это связано с тем, что генеративные деревья ели находятся на значительном удалении от рассматриваемого сосняка-зеленомошника. В этом сообществе малая численность теневыносливой ели не мешает развитию ценопопуляций светолюбивого дуба.
Этап II–Д. Сосняк-зеленомошник с преобладанием im и v подроста дуба (рис. 10). Ярус древостоя представлен культурами сосны, возраст которых составляет 80 лет. Последний низовой пожар был 35 лет назад. Онтогенетическая структура ценопопуляции сосны – прерывисто неполночленная. Во взрослой фракции господствуют g1 и g2 деревья, а в подросте, по сравнению с предыдущим этапом, сохранились только единичные im особи низкой жизненности (рис. 9, II).

Рисунок 10. Сосняк-зеленомошник с преобладанием im и v подроста дуба (этап II–Д)
Обозначения: А – общий вид сообщества; Б – виргинильный подрост дуба первой подгруппы (v1). Дата фотографий – июнь 2024 г.
В сообществе продолжает развиваться инвазионная ценопопуляция дуба. Численность подроста дуба продолжает возрастать, в ней появились v особи (рис. 9, II). В ценозе также присутствует ель, однако численность ее ценопопуляции остается на низком уровне. На одном гектаре встречено только 23 особи, представленные im и v подростом. Такие ценопопуляции называют фрагментарными. Другими словами, дуб в этом сообществе может нормально развиваться, поскольку отсутствует конкуренция со стороны подроста ели. Единичный подрост груши, берез бородавчатой и пушистой, встреченный в сообществе, также не оказывает существенного влияния на развитие ценопопуляций дуба.
Этап III–Д. Сосняк-зеленомошник с преобладанием im–g1 особей дуба (рис. 11). Ярус древостоя представлен культурами сосны, возраст которых составляет 115 лет. В сообществе последний низовой пожар был 70 лет назад. Структура ценопопуляции сосны – прерывисто неполночленная. Во взрослой фракции господствуют только g2 деревья, а в подросте найдено небольшое число j и im особей низкой жизненности (рис. 9, III). В древостое представлена также береза бородавчатая. Ее ценопопуляция прерывисто неполночленная, она сформирована небольшим числом особей (рис. 12, III).

Рисунок 11. Сосняк-зеленомошник с преобладанием im–g1 особей дуба (этап III–Д)
Обозначения: А – общий вид сообщества; Б – молодой генеративный дуб (g1). Даты фотографий: А – октябрь 2024 г.; Б – начало марта 2025 г.
У дуба за 70 лет отсутствия пожаров сформировалась левосторонняя онтогенетическая структура ценопопуляции, в ней отсутствуют только g3 и s особи (рис. 9, III). На предыдущих этапах развития сообществ ценопопуляции дуба формировались только за счет притока желудей из окружающих сообществ. В этом ценозе у дуба появились собственные источники семян – g1 и g2 деревья. В результате численность дуба возросла почти в два раза. Левосторонняя онтогенетическая структура и высокая численность свидетельствует о том, что у дуба сформировался устойчивый оборот поколений. Возникновение устойчивой (дефинитивной) структуры популяции дуба определяется малым участием в сообществе теневыносливой ели – основного конкурента дуба в сосняках на песчаных почвах. Численность ели небольшая, всего 30 особей на га. Однако в ее ценопопуляции появились единичные g1 растения: ель с этого этапа развития начинает сама поддерживать свою численность (рис. 9, III). Следует отметить, что в структуре рассматриваемого сосняка сформировалась инвазионная ценопопуляция клена остролистного с малым числом особей (рис. 12, III). Известно, что этот теневыносливый вид – один из конкурентов дуба (Погребняк, 1968; Атрохин, 1982). В травяном покрове этого и следующего сообщества появляются единичные im особи яблони лесной, их ценопопуляции фрагментарного типа.
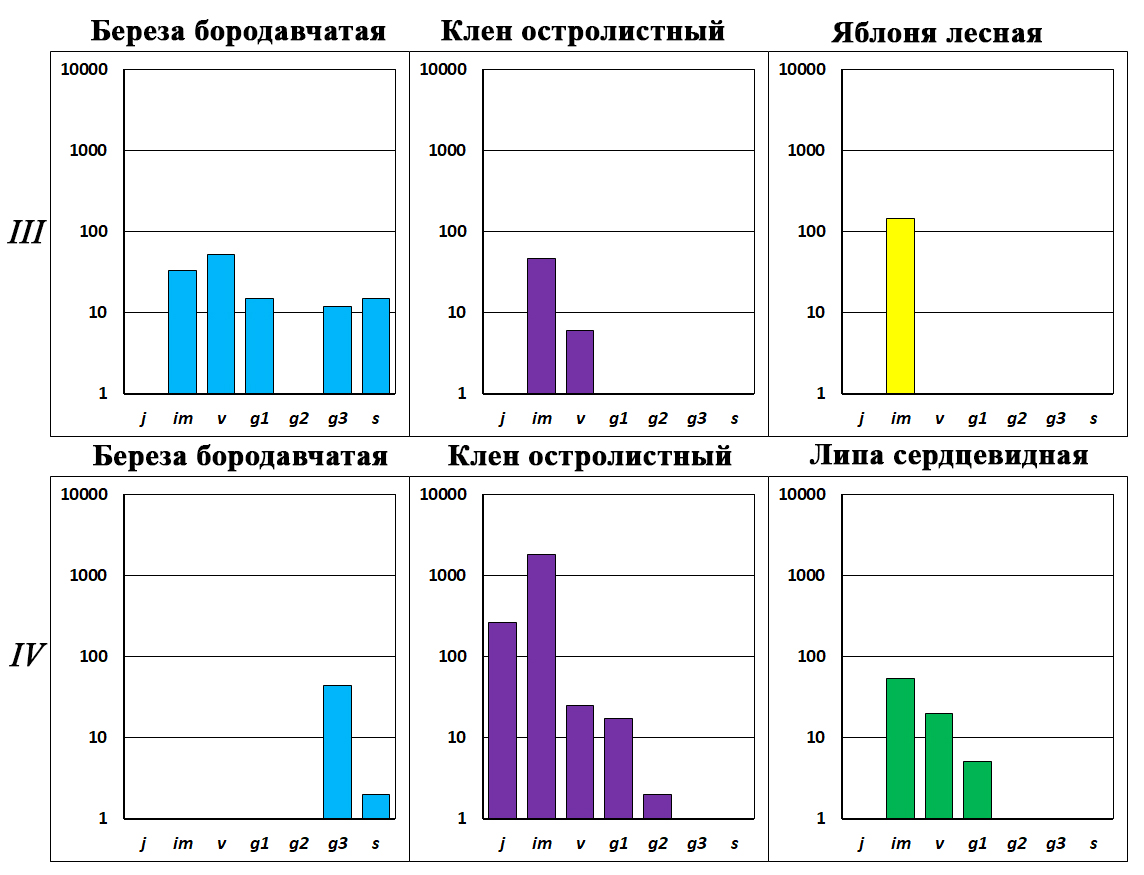
Рисунок 12. Онтогенетический состав ценопопуляций деревьев в сосняках-зеленомошниках, которые расположены вблизи плодоносящих дубов черешчатых и вдали от генеративных елей обыкновенных и других широколиственных деревьев (продолжение)
Обозначения. Римские цифры – этапы формирования ценопопуляций (см. текст). По оси х – онтогенетические состояния, по оси y – число особей на 1 га в логарифмическом масштабе
Этап IV–Д. Хвойно-широколиственный лес с g2–s дубом (рис. 13). В ярусе древостоя доминируют два вида – сосна обыкновенная и дуб черешчатый. Возраст сосны – 95 лет. В сообществе не обнаружены следы пожаров. Следует отметить, что в ярусе древостоя появляется большое число особей ели и клена, а также единичные молодые генеративные деревья липы. Это свидетельствует о том, что в ходе сукцессии меняется тип леса: сосняк-зеленомошник преобразуется в сообщества хвойно-широколиственного леса.
Структура ценопопуляции сосны относится к регрессивному типу, в ней присутствуют только g2 и g3 особи, а подрост полностью отсутствует (рис. 9, IV). Число генеративных деревьев сосны по сравнению с предыдущими этапами уменьшилось в семь–десять раз. Это связано с выходом в ярус древостоя дуба, ели и клена. Они затеняют крону светолюбивой сосны, которая начинает преждевременно стареть и вываливаться из сообщества. В ярусе древостоя присутствует береза бородавчатая, ее ценопопуляция регрессивного типа, в ней присутствуют только g3 и s деревья (рис. 12, IV). Через два–три десятилетия береза полностью вывалится из сообщества.

Рисунок 13. Хвойно-широколиственный лес с g2–s дубом (этап IV–Д)
Обозначения: А – общий вид сообщества; Б – зрелый генеративный дуб (g2) аэроксильной жизненной формы; В – погибший взрослый дуб. Даты фотографий: А, Б – октябрь 2024 г.; В – начало марта 2025 г.
Возрастная структура дуба полночленная, в ней представлены растения всех онтогенетических состояний – от j до g3 и s особей (рис. 9, IV). Несмотря на это ценопопуляция дуба начинает деградировать. Так, в ее структуре существенно увеличивается доля g3 и s особей. Известно, что плодоношение у g3 растений ослаблено, а у s – отсутствует (Евстигнеев, Короткова, 2024). В результате значительно сокращается участие j, im, v и g1 растений. Их численность по сравнению с предыдущим этапом уменьшается в 15 раз. В результате молодых особей дуба становится недостаточно для обеспечения устойчивого оборота поколений. Причина такой деградации дуба – появление в сообществе генеративных елей, кленов и лип. Они затеняют крону дуба: его фотосинтетическая поверхность сокращается, дышащие части растения (корни, стволы, ветви) получают меньше пластических веществ, деревья начинают быстро стареть, а затем засыхают, ломаются или вываливаются (рис. 13, В). Одновременно ель, клен и липа, которые сформировали сомкнутый полог во всех ярусах сообщества (рис. 12, IV), затеняют и вытесняют светолюбивый подрост дуба, который периодически появляется в травяном покрове.
Ценопопуляционный анализ сосняков-зеленомошников, которые расположены рядом с плодоносящими дубами и вдали от генеративных особей ели и других широколиственных деревьев (клена, липы), выявил некоторые особенности формирования ценопопуляций дуба. В этих условиях дуб практически сразу после низовых пожаров начинает внедряться в сосняки. Сначала он формирует ценопопуляции инвазионного типа с высокой численностью особей, а затем к 70 годам – ценопопуляции левостороннего типа с устойчивый оборотом поколений. Однако через 100 лет в этих сообществах появляются теневыносливые ель, клен и липа. Для ценопопуляций клена и ели характерны высокая численность особей и непрерывный оборот поколений. Они со временем полностью вытеснят из сообществ светолюбивые деревья – дуб и сосну. Все это свидетельствует о том, что в ходе сукцессионных преобразований на месте сосняка-зеленомошника постепенно сформируется субклимаксный олигодоминантный елово-широколиственный лес без участия сосны и дуба.
ЗАКЛЮЧЕНИЕ
Доминант большинства сообществ на песчаных почвах – сосна обыкновенная. При естественном развитии ценозов сосна постепенно исчезнет, а сообщества преобразуются в ельники-зеленомошники или в хвойно-широколиственные леса. Это связано с чрезвычайно слабой теневыносливостью сосны (Евстигнеев, 2022), которая не позволяет ей возобновляться в сомкнутых лесах. В этих сообществах с течением времени формируется регрессивная или регрессивно-инвазионная онтогенетическая структура ее популяций, при которой невозможен устойчивый оборот поколений: генеративные особи стареют и вываливаются, а появляющийся подрост гибнет в j или im состояниях от недостатка света. Одна из причин деградации ценопопуляций сосны – это внедрение в сообщество более теневыносливых видов деревьев (ели, дуба, клена, липы).
В работе выявлено два основных направления сукцессионных преобразований сосняков-зеленомошников. Пути изменения сосняков определяются дальностью расположения генеративных деревьев дуба, ели, клена и липы.
В случае близкого расположения генеративных деревьев ели и дуба доминантом во всех ярусах сосняков-зеленомошников постепенно становится ель обыкновенная. Беспрепятственное развитие популяций ели связано с ее высокой теневыносливостью и большой скоростью роста по высоте, а также с отсутствием заноса семян конкурентов: липы и клена. Эти виды могли бы существенно ограничить участие ели в сообществах: их теневыносливость и скорость роста по высоте сопоставимы с елью (Евстигнеев, 2004, 2022). Однако плодоносящие широколиственные деревья расположены в двух километрах от рассматриваемых сообществ. И. Ф. Удра (1988) показал, что в лесном покрове скорость расселения липы составляет 3 м в год, а клена – 5 м. При этих обстоятельствах липа может вселиться в сообщества только спустя 600–700 лет, а клен – не раньше 400 лет. В таких сосняках благодаря сойкам в течение первых сорока–пятидесяти лет активно формируются инвазионные ценопопуляции дуба с высокой численностью особей. Однако теневыносливая ель, обладая большей скоростью роста, после пятидесяти лет обгоняет светолюбивый дуб по высоте, затеняет его и вытесняет из сообщества. Таким образом, сосняки-зеленомошники в непосредственной близости от генеративных деревьев дуба и ели сменяются сначала олигодоминантными елово-сосновыми лесами. После старения и выпадения сосны сообщества преобразуются в субклимаксные монодоминантные разновозрастные ельники-зеленомошники с устойчивым оборотом поколений в ценопопуляциях ели.
Сукцессионные преобразования сосняков-зеленомошников, которые расположены в непосредственной вблизи от плодоносящих дубов и вдали от источников диаспор ели, липы и клена (300 м и далее), приводят к формированию хвойно-широколиственных лесов с большим участием дуба. Однако ценопопуляционный анализ свидетельствует о том, что в рассматриваемых сообществах только первое поколение дуба способно пройти полный онтогенез. Последующие поколения дуба, появляющиеся в сообществе, погибают на ранних этапах развития из-за внедрившихся популяций теневыносливых деревьев – ели, клена, липы. Эти виды обгоняют подрост дуба в развитии, затеняют его и вытесняют из сообщества. Исследование показало, что ель начала активно внедряться в сосняки-зеленомошники спустя 40 лет после низового пожара, клен – через 70 лет, а липа – по истечении 100 лет. Это время, которое необходимо этим видам для преодоления расстояния в 300 м. Оно практически полностью согласуется с цифрами по скорости расселения ели, клена и липы, которые приводит И. Ф. Удра (1988). Такое отложенное внедрение ели, клена и липы в сообщество позволяет первому поколению дуба стать эдификатором хвойно-широколиственных лесов. После выпадения из ценоза светолюбивых сосны и дуба, которые не способны возобновляться в лесах теневого типа, в древостое и подросте абсолютное доминирование получают теневыносливые виды – ель, клен и липа. Со временем ценопопуляции этих деревьев сформируют субклимаксные олигодоминантные разновозрастные елово-широколиственные леса с устойчивым оборотом поколений без участия дуба и сосны.
Формирование разновозрастных монодоминантных ельников и олигодоминантных елово-широколиственных лесов возможно только при абсолютном заповедании. За пределами заповедника частые низовые пожары, провоцируемые человеком, время от времени уничтожают внедряющиеся виды растений и приостанавливают восстановление лесных сообществ на многие десятилетия.
ФИНАНСИРОВАНИЕ
Исследование выполнено в рамках молодежной лаборатории ЦЭПЛ РАН «Биоразнообразие и климаторегулирующие функции лесов» (регистрационный номер 1024100700084-5-1.6.19)
СПИСОК ЛИТЕРАТУРЫ
Атрохин В. Г. Лесоводство и дендрология. М.: Лесн. пром-сть, 1982. 368 с.
Браславская Т. Ю. Популяционная организация лесообразующих видов в пойме равнинной средней реки (на примере заповедника «Большая Кокшага»). М.: Цифровичок, 2019. 112 с.
Восточноевропейские леса: история в голоцене и современность. М.: Наука, 2004а. Кн. 1. 479 с.
Восточноевропейские леса: история в голоцене и современность. М.: Наука, 2004б. Кн. 2. 575 с.
Восточноевропейские широколиственные леса. М.: Наука, 1994. 364 с.
Гроздов Б. В. Типы леса Брянской, Смоленской и Калужской областей. Брянск: Брянский лесохозяйственный ин-т, 1950. 56 с.
Евстигнеев О. И. Механизмы поддержания биологического разнообразия лесных биогеоценозов. Дис. … док. биол. наук. Нижний Новгород: НГУ им. Н.И. Лобачевского, 2010. 513 с.
Евстигнеев О. И. Неруссо-Деснянское полесье: история природопользования. Брянск: группа компаний Десяточка, 2009. 139 с.
Евстигнеев О. И. Отношение деревьев к свету: онтогенетический аспект. М.: Перо, 2022. 36 с.
Евстигнеев О. И. Поливариантность сосны обыкновенной в Брянском полесье // Лесоведение. 2014. № 2. С. 69–77.
Евстигнеев О. И. Популяционные стратегии видов деревьев // Восточноевропейские леса: история в голоцене и современность. М.: Наука, 2004. Кн. 2. С. 176–205.
Евстигнеев О. И., Воеводин П. В. Видовой состав сосняка-зеленомошника на разном удалении от хвойно-широколиственного леса // Вестник Брянского гос. ун-та. Серия точные и естественные науки. Брянск: РИО БГУ, 2012. № 4. С. 74–76.
Евстигнеев О. И., Воеводин П. В., Коротков В. Н., Мурашев И. А. Зоохория и дальность разноса семян в хвойно-широколиственных лесах Восточной Европы // Успехи соврем. биол. 2013. Т. 133. № 4. С. 392–400.
Евстигнеев О. И., Коротков В. Н., Беляков К. В., Браславская Т. Ю., Романовский А. М., Рубашко Г. Е., Сарычева Е. П., Федотов Ю. П. Биогеоценотический покров Неруссо-Деснянского полесья: механизмы поддержания биологического разнообразия. Брянск: Заповедник Брянский лес, 1999. 176 с.
Евстигнеев О. И., Короткова Н. В. Поливариантность онтогенеза дуба черешчатого в сосняках Брянского полесья // Russian Journal of Ecosystem Ecology. 2024. Vol. 9. No. 1. P. 1–31.
Евстигнеев О. И., Мурашев И. А., Коротков В. Н. Анемохория и дальность рассеивания семян деревьев Восточноевропейских лесов // Лесоведение. 2017. № 1. С. 45–52.
Загреев В. В., Сухих В. И., Швиденко А. З., Гусев Н. Н., Мошкалев А. Г. Общесоюзные нормативы для таксации лесов. М., 1992. 495 с.
Заугольнова Л. Б., Жукова Л. А., Комаров А. С., Смирнова О. В. Ценопопуляции растений (очерки популяционной биологии). М.: Наука, 1988. 184 с.
Кикеева А. В., Новичонок Е. В., Софронова И. Н., Крышень А. М. Особенности раннего развития сеянцев Picea abies (Pinaceae) на ксилолитическом субстрате // Ботанический журнал. 2022. Т. 107. № 10. С. 996–1009.
Коротков В. Н. Демутационные процессы в островных лесных массивах (на примере ГИЗЛ «Горки Ленинские» и Каневского заповедника). Автореф. … канд. дис. М., 1992. 16 с.
Корчагин А. А. Определение возраста деревьев умеренных широт // Полевая геоботаника. М.-Л.: Наука, 1960. Т. 2. С. 209–240.
Кулешова Л. В., Коротков В. Н., Потапова Н. А., Евстигнеев О. И., Козленко А. Б., Русанова О. М. Комплексный анализ послепожарных сукцессий в лесах Костомукшского заповедника (Карелия) // Бюл. МОИП. Отд. биол. 1996. Т. 101. Вып. 4. С. 3–15.
Мелехов И. С. Лесоводство. М.: Агропромиздат, 1989. 301 с.
Мелехов И. С. О взаимоотношениях между сосною и елью в связи с пожарами в лесах европейского Севера СССР // Ботанический журнал. 1944. Т. 29. № 4. С 131–135.
Морозов Г. Ф. Учение о лесе. М.-Л.: Гослесбумиздата, 1949. 456 с.
Морозова О. В. Леса заповедника «Брянский лес» и Неруссо-Деснянского полесья (синтаксономическая характеристика). Брянск: Заповедник Брянский лес, 1999. 98 с.
Панченко С. М. Лесная растительность национального природного парка «Деснянско-Старогутский». Сумы: Университетская книга, 2013. 311 с.
Погребняк П. С. Общее лесоводство. М.: Колос, 1968. 440 с.
Растительность европейской части СССР. Л.: Наука, 1980. 429 с.
Ремезов Н. П. Погребняк П. С. Лесное почвоведение. М.: Лесн. пром-сть, 1965. 324 с.
Романовский А. М. Поливариантность онтогенеза Picea abies (Pinaceae) в Брянском полесье // Ботанический журнал. 2001. Т. 86, № 8. С. 72–85.
Санников С. Н., Санникова Н. С. Экология естественного возобновления сосны под пологом леса. М.: Наука, 1985. 152 с.
Смирнова О. В., Возняк Р. Р., Евстигнеев О. И., Коротков В. Н., Носач Н. Я., Попадюк Р. В., Самойленко В. К., Торопова Н. А. Популяционная диагностика и прогнозы развития заповедных лесных массивов (на примере Каневского заповедника) // Ботанический журнал. 1991. Т. 76. № 6. С. 860–871.
Смирнова О. В., Коротков В. Н. Старовозрастные леса Пяозерского лесхоза северо-западной Карелии // Ботанический журнал. 2001. Т. 86. № 1. С. 98–109.
Смирнова О. В., Торопова Н. А. Сукцессия и климакс как экосистемный процесс // Успехи соврем. биол. 2008. Т. 128. № 2. С. 129–144.
Смирнова О. В., Чистякова А. А., Дробышева Т. И. Ценопопуляционный анализ и прогнозы развития дубово-грабовых лесов Украины // Журнал общей биологии. 1987. Т. 48. № 2. С. 200–212.
Смирнова О. В., Чистякова А. А., Рипа С. И., Лысых Н. И. Популяционная организация буковых лесов Закарпатья // Бюл. МОИП. Отд. биол. 1989. Т. 94. Вып. 5. С. 78–91.
Сукачев В. Н. Дендрология с основами лесной геоботаники. Л.: Гослестехиздат, 1938. 575 с.
Ткаченко М. Е. Общее лесоводство. М.–Л.: Гослесбумиздат, 1952. 600 с.
Удра И. Ф. Расселение растений и вопросы палео- и биогеографии. Киев: Наукова Думка, 1988. 197 с.
Цветков М. А. Изменение лесистости Европейской России с конца XVII столетия по 1914 год. М.: Изд-во АН СССР, 1957. 202 с.
Чистякова А. А. Большой жизненный цикл Tilia cordata Mill. // Бюл. МОИП. Отд. биол. 1979. Т. 84. Вып. 1. С. 85–98.
Юркевич И. Д., Ловчий Н. Ф., Гельтман Е. С. Леса Белорусского Полесья (геоботанические исследования). Минск: Наука и техника, 1977. 288 с.
Evstigneev O. I. Ontogenetic scales of relation of trees to light (on the example of Eastern European forests) // Russian Journal of Ecosystem Ecology. 2018. Vol. 3. No. 3. P. 1–18.
Evstigneev O. I., Korotkov V. N. Ontogenetic stages of trees: an overview // Russian Journal of Ecosystem Ecology. 2016а. Vol. 1. No. 2. P. 1–31.
Evstigneev O. I., Korotkov V. N. Pine forest succession on sandy ridges within outwash plain (sandur) in Nerussa-Desna Polesie // Russian Journal of Ecosystem Ecology. 2016б. Vol. 1. No. 3. P. 1–18.
Smirnova O. V., Bobrovsky M. V., Braslavskaya T. Yu, Evstigneev O. I., Khanina L. G., Korotkov V. N. et al. European Russian Forests. Their Current State and Features of Their History. Springer Heidelberg. 2017. 566 p.
Рецензент: д.б.н., доцент Семенищенков Ю. А.





