- DOI: 10.31509/2658-607x-202583-176
- УДК 581.526.42:004.942
Анализ методами имитационного моделирования реакции ценопопуляций черники обыкновенной на внешние воздействия
![]()
© 2025 П. В. Фролов1*, Е. В. Зубкова1, В. Н. Шанин1,2,3
1Институт физико-химических и биологических проблем почвоведения РАН – обособленное подразделение Федерального исследовательского центра «Пущинский научный центр биологических исследований Российской академии наук»
Россия, 142290 Московская обл., г. Пущино, ул. Институтская, д. 2, корп. 2.
2 Центр по проблемам экологии и продуктивности лесов им. А. С. Исаева Российской академии наук
Россия, 117997 Москва, ул. Профсоюзная, 84/32, стр. 14
3Институт математических проблем биологии РАН – филиал Института прикладной математики им. М. В. Келдыша РАН
Россия, 142290 Московская обл., г. Пущино, ул. проф. Виткевича, д. 1
*E-mail: ximikadze@gmail.com
Поступила в редакцию: 20.05.2025
После рецензирования: 15.09.2025
Принята к печати: 21.09.2025
Растения травяно-кустарничкового яруса вносят существенный вклад в биологический круговорот в лесных экосистемах. Однако количественные реакции данного компонента на нарушающие воздействия различного характера, а также его восстановление после повреждений, до сих пор остаются мало изученными. В статье представлены результаты исследования реакции ценопопуляций черники обыкновенной (Vaccinium myrtillus L., 1753) на различные типы нарушающих воздействий. Полевые эксперименты проводились в сосняке чернично-ландышево-зеленомошном (Московская область) и включали три типа воздействий: удаление подстилки, срезание побегов и перерубание корневищ. Результаты показали, что при повреждении побегов или корневищ выживаемость парциальных кустов зависит от сохранения физиологической интеграции с клоном. Основным механизмом восстановления является пробуждение спящих почек на корневищах. Удаление подстилки приводит к ортотропному росту корневищ и формированию новых побегов. С использованием этих данных был разработан и добавлен в модель CAMPUS‑S модуль внешних воздействий, позволяющий имитировать влияние пожаров и рубок на ценопопуляции черники, а также восстановление кустарничков после подобных нарушений. Имитационные эксперименты выявили, что ценопопуляции черники устойчивы к разовым повреждениям, но систематические воздействия приводят к их деградации. Низовые пожары со средней способностью к распространению (до 10% территории) не вызывают гибели популяции, но при 15% повреждении восстановление невозможно. Выборочные рубки, особенно с применением гусеничной техники, замедляют восстановление, но не приводят к гибели ценопопуляций черники. Результаты могут служить для оценки устойчивости ценопопуляций и позволят планировать систему лесохозяйственных мероприятий с точки зрения минимизации ущерба для травяно-кустарничкового яруса.
Ключевые слова: Vaccinium myrtillus, имитационное моделирование, CAMPUS‑S, пожары, рубки леса, устойчивость ценопопуляций, лесные экосистемы, сосняки
Кустарнички рода Vaccinium, в т.ч. черника обыкновенная, брусника обыкновенная и др., широко распространены в таёжных лесах, лесотундре и тундре. Они вносят значительный вклад в недревесную продукцию в лесных экосистемах (Бобкова, 1987; Morén, Lindroth, 2000) и участвуют в биогеохимическом круговороте элементов, играя заметную роль в восстановлении лесов после сильных внешних воздействий (Tolvanen, 1994).
Ценопопуляции черники могут выдерживать различные внешние воздействия, такие как вытаптывание (Emanuelsson, 1984), выедание (Ericson, 1977; Emanuelsson, 1984; Laine, 1988; Atlegrim, 1991, Dahlgren et al., 2007), промерзание почвы (Havas, 1971; Sjörs, 1989), а также выборочные рубки (Валова, 1973). Одной из важных причин устойчивости клонов черники является наличие запасов углерода и питательных веществ, большая часть которых хранится в подземных побегах – корневищах (Flower‑Ellis, 1971; Pakonen et al., 1991) и может быть мобилизована в случае повреждения отдельных парциальных образований (Dahlgren et al., 2007; Маслова, 2014). Основным механизмом восстановления клонов черники после повреждения является активация спящих почек, расположенных на корневищах (Emanuelsson, 1984).
Такой механизм восстановления особенно важен в местообитаниях северных широт, которые характеризуются коротким вегетационным периодом и дефицитом питательных веществ, особенно азота, в почве. Эти факторы ограничивают рост и восстановление ценопопуляций растений после повреждения (Archer, Tieszen, 1980; Haag, 1974; Mattson, 1980; Cargill, Jeffries, 1984).
В начале 2000-х годов были разработаны и апробированы индивидуально-ориентированные пространственно-детализованные имитационные модели, позволяющие оценить параметры восстановления и самоподдержания ценопопуляций длиннокорневищных растений при различных нарушениях (Михайлова и др., 2008). Эти модели изначально разрабатывались как классические клеточно-автоматные, но в дальнейшем их структура была усложнена. Они были названы решётчатыми, так как в таких моделях пространство, на котором имитируется популяционная динамика, задано решёткой (двумерным массивом квадратных ячеек с целочисленными координатами), что позволяет упростить некоторые вычислительные алгоритмы. В этих работах было показано, что скорость захвата территории длиннокорневищными видами травянистых растений зависит от длительности онтогенетических состояний, а также от параметров вегетативного разрастания и интенсивности семенного размножения. Вычислительные эксперименты показывают, что, например, в отсутствие конкуренции за свободную территорию ценопопуляция Stellaria holostea L. (1753) осуществляет самоподдержание в основном вегетативным способом, ценопопуляция Asarum europaeum L. (1753) – семенным, а ценопопуляция Aegopodium podagraria L. (1753) – смешанным (Михайлова и др., 2008). Было подтверждено ранее постулировавшееся (Комаров, 1982; Комаров, Паленова, 2002) существование пороговых значений доли непригодных микроучастков, выше которых самоподдержание популяций прекращается. Такие работы были позже повторены для других видов венгерскими исследователями (Oborny, Englert, 2012).
В последние годы накоплен значительный объём экспериментальных данных, раскрывающих структуру популяций растений рода Vaccinium, влияние внешних условий (богатство почвы, освещённость, водный режим) на их продуктивность, охарактеризованы морфологические особенности кустарничков, оценён потенциал урожайности при различных сценариях ведения лесного хозяйства (Колычева, Чумаченко, 2022). Однако количественная динамика их ценопопуляций при различных повреждениях до сих пор остаётся неизученной.
Целью данного исследования явилось выявление при помощи методов имитационного моделирования максимально допустимой интенсивности повреждений различного характера: рубок, пожаров (как разовых, так и систематических), при которых ценопопуляции черники могут восстанавливаться.
МАТЕРИАЛЫ И МЕТОДЫ
Полевые эксперименты. Полевой эксперимент по выявлению реакции клонов черники на различные нарушающие воздействия проводился в сосняке чернично-ландышево-зеленомошном (Московская область, ГКУ МО Мособллес, филиал Русский лес, Данковское участковое лесничество).
В ходе полевого эксперимента было заложено 9 пробных площадей (ПП) размером от 16 до 25 м2 каждая (в зависимости от размеров клона черники), границы которых определялись границей распространения надземных побегов клона (рис. 1А). Характеристики местообитаний, которые фиксировались при описании пробных площадей, приведены в таблице 1.
Таблица 1. Обобщённые характеристики пробных площадей
| Характеристика | Значение |
| Микрорельеф | Выражен |
| Условия увлажнения | Хорошо дренированная почва |
| Почва | Серая лесная (на песках) |
| Влияние животных | Порои кабанов |
| Окружение | Приспевающий сосняк с подростом дуба черешчатого (Quercus robur L. (1753)), березы (Betula spp.), сосны обыкновенной (Pinus sylvestris L. (1753)), с подлеском из рябины обыкновенной (Sorbus aucuparia L. (1753)) и крушины ломкой (Frangula alnus Mill. (1768)) |
| Хозяйственное использование | Рекреация |
| Мёртвый покров | Выражен |
| Аспект | Зелёный |
| Общее проективное покрытие травяно-кустарничкового яруса | 65–70% |
| Сомкнутость крон | 0.6–0.7 |
| Формула древостоя | 10С |
| Ассоциация | Сосняк чернично-ландышево-зеленомошный |
В пределах каждой ПП были выделены по 4 площадки размером 0.25 м2 (0.5 × 0.5 м) с побегами черники, расположенные на максимальном удалении друг от друга (рис. 1Б). На трёх из четырёх малых площадок в пределах каждой ПП был осуществлён один из видов нарушающих воздействий: (1) убрана подстилка; (2) срезаны парциальные кусты на уровне почвы; (3) перерублены корневища (крест-накрест по диагоналям малой площадки). Четвёртая малая площадка являлась контролем.
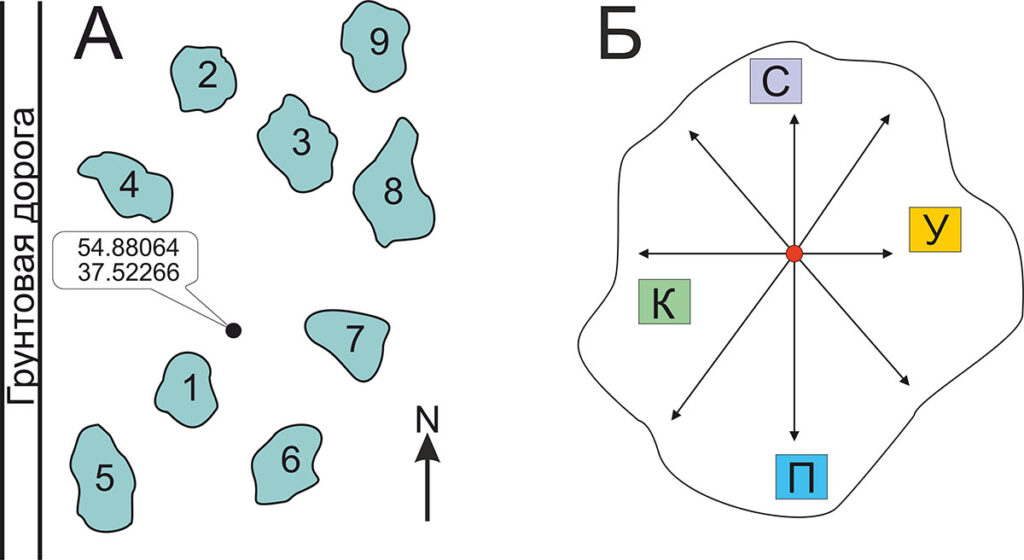
Рисунок 1. А – Схема расположения и форма пробных площадей. Размеры пробных площадей и расстояния между ними изображены в разном масштабе. Б – пример расположения малых площадок на ПП: К – контроль, С – срезаны парциальные кусты, У – убрана подстилка, П – перерублены корневища. Линия – границы клона, ● – центр клона
Наблюдения и описания площадок проводились через 1, 2, 4, 8 и 12 недель после закладки эксперимента. При наблюдении через 12 недель на всех площадках был убран моховой покров и зафиксировано количество и относительное расположение парциальных кустов, сформировавшихся с момента начала эксперимента. При промежуточных наблюдениях (через 1, 2, 4 и 8 недель после закладки эксперимента) использовались только неинвазивные (во избежание дополнительных повреждений клона) методы – фиксировалось количество отмерших парциальных кустов, а также их фенофазы. Наблюдение за площадками, на которых удалялась лесная подстилка, было продолжено на следующий год (через месяц после начала вегетации).
Для сравнения распределений выживших, погибших и новых парциальных кустов при различных типах воздействий применялся непараметрический Q‑тест Кохрана и парный тест Макнемара с поправкой Бонферрони (Stephen, Adruce, 2018). Для выявления зависимости количества новых парциальных образований от количества отмерших при перерубании корневищ и срезании побеговой системы был проведён регрессионный анализ в среде статистического программирования R версии 4.2.2 (R Core Team, 2022) методом нелинейной регрессии.
Имитационные эксперименты. Модель CAMPUS‑S (Frolov et al., 2020a,b) была разработана для моделирования ценопопуляций растений травяно-кустарничкового яруса (ТКЯ) в лесных экосистемах и их вклада в круговорот углерода и азота. Модель использует несколько методов моделирования: технику L‑систем и клеточных автоматов, а также матричное моделирование. Видоспецифичные особенности определяются для каждой онтогенетической стадии (L-системы), но их реализация для конкретных особей в процессе моделирования зависит от внешних условий (клеточные автоматы). Для учёта вариативности переходов между онтогенетическими стадиями применяется метод матричного моделирования. Пространственное разрешение модели составляет 1 см2, а временной шаг – один месяц. Модель состоит из двух подмоделей: подмодели структуры растительных популяций и подмодели динамики углерода и азота в растениях и почве. В модели CAMPUS‑S пространство неоднородно. Карта микрорельефа динамически загружается из внешнего источника, и на её основе в процессе моделирования генерируется пространственная неоднородность температуры и влажности почвы. Для реализации механизма обратной связи между растениями и почвой выполнена интеграция с моделью динамики органического вещества почвы Romul_Hum (Komarov et al., 2017; Chertov et al., 2017a,b), которая описывает трансформацию растительного опада и разных фракций органического вещества почвы в зависимости от их химических свойств и гидротермических условий, а одной из выходных переменных модели является количество азота в почве в доступных для растений формах.
Модификация модели CAMPUS‑S. На основе данных, полученных в результате полевого эксперимента, а также литературных данных, модель CAMPUS‑S была дополнена модулем моделирования внешних воздействий, состоящим из блока имитации внешних нарушающих воздействий и блока ввода вариантов реакции на внешние воздействия.
Блок имитации внешних нарушающих воздействий позволяет задать как отдельные виды нарушений (удаление подстилки, уничтожение побеговой системы, перерубание корневищ), так и их сочетания, имитирующие реальные сценарии естественных нарушений и повреждений, связанных с деятельностью человека. Низовые лесные пожары могут имитироваться посредством имитации уничтожения подстилки и побеговой системы растений ТКЯ. Для имитации влияния выборочных рубок в модели воспроизводится уничтожение подстилки и побеговой системы, а также перерубание корневищ. Более подробное описание имитационных сценариев приведено ниже.
В случае выбора отдельных воздействий задаётся их масштаб (доля нарушаемой территории), а также либо конкретный шаг моделирования, на котором будет произведено воздействие, либо периодичность данного воздействия. Распределение нарушаемых областей в пространстве имитационной площадки в данном случае происходит случайным образом (здесь и далее под случайным распределением подразумевается псевдослучайное распределение, полученное при помощи виткового регистра сдвига с обобщённой отдачей (вихрь Мерсенна) в реализации алгоритма MT19937 (Matsumoto, Nishimura, 1998)).
В случае выбора комплексных воздействий, помимо масштаба и периодичности воздействия, реализована возможность задания ряда специфичных параметров.
Имитация низовых лесных пожаров. При имитации низовых лесных пожаров учитывается тот факт, что характер и степень повреждения определённой территории зависит от её микрорельефа и, соответственно, влажности (Комарова, 1993). Точка возникновения пожара выбирается случайным образом. Далее происходит выбор ячеек на решётке, на которых возможно круговое распространение огня (с центром в ячейке возникновения пожара). При этом учитывается способность к распространению пожара, которая определяется как пороговое значение влажности подстилки в ячейке, выше которого распространение пожара в данную ячейку невозможно. В случае если n ячеек подряд оказываются «устойчивыми к возгоранию» (влажность в ячейке оказывается выше способности пожара к распространению), распространение огня в данном направлении прекращается. Величина значения n вводится пользователем и позволяет имитировать различную скорость ветра, как главный фактор распространения пожара.
Имитация рубок. При моделировании рубок задается площадь территории, на которой происходит вырубка, шаг модели, на котором происходит рубка, а также применяемая техника (на шинах низкого давления или на гусеничном ходу).
Вероятность повреждения ячейки в ходе рубки рассчитывается согласно уравнению:
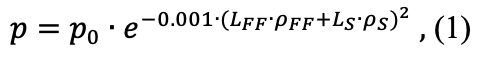
где p – вероятность повреждения ячейки, p0 – вероятность повреждения напочвенного покрова при отсутствии подстилки и снежного покрова в зависимости от способа рубки, LFF и LS – мощность органогенного горизонта почвы и толщина снежного покрова, соответственно, ρFF и ρS – плотность органогенного горизонта почвы и снега, соответственно (кг м−3). Рассматривались крайние по степени воздействия на живой напочвенный покров варианты. В качестве наиболее мягкого варианта рассматривалась техника на шинах низкого давления (ШНД), при использовании которой величина p0 составляет 0.4 (Бойчук, Дроздов, 1991). В качестве экстремального варианта рассматривалось использование валочной машины ВМ-4 на гусеничном шасси (ГШ) с последующей трелёвкой. У данной машины в процессе эксплуатации выявили целый ряд существенных недостатков, в частности отсутствие гидроманипулятора. Вследствие этого, узкозахватность вынуждала оператора машины каждый рабочий цикл начинать с подъезда к каждому дереву, что делало практически невозможным сохранение подроста и напочвенного покрова. При использовании данной машины p0 = 0.9 (Рожин, Григорьев, 1972). График функции, используемой при моделировании зависимости вероятности повреждения ячейки, представлен на рисунке 2.
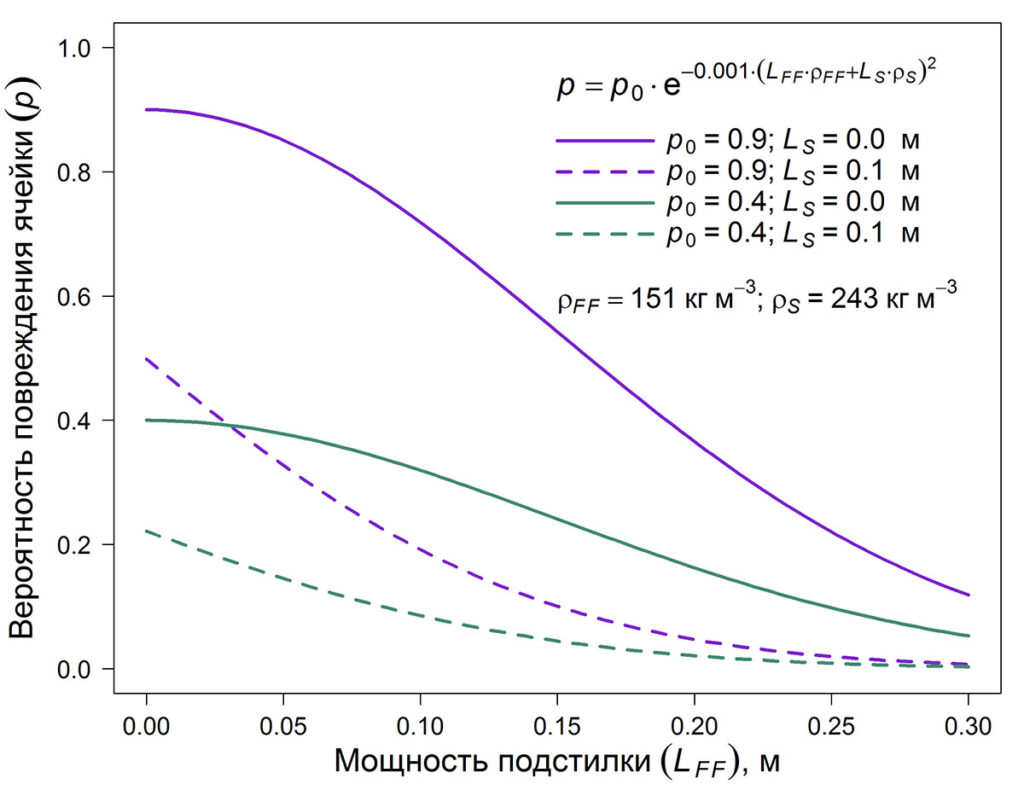
Рисунок 2. Моделируемая зависимость вероятности повреждения ячейки от способа рубки, мощности и плотности подстилки и снежного покрова
В квадрате площадью, равной площади территории, на которой производится вырубка, вероятность повреждения каждой ячейки равняется p. Моделирование территории, повреждаемой с вероятностью 100%, происходит в соответствии со схемой разработки лесосек с сохранением подроста методом узких пасек (рис. 3) (Мариничев, Мариничева, 2016). При этой схеме лесосека разбивается на пасеки шириной 35 м. Пасечные волоки шириной 4 м прокладываются посередине. Погрузочная площадка размером 40 × 40 м. располагается в одном из углов лесосеки.
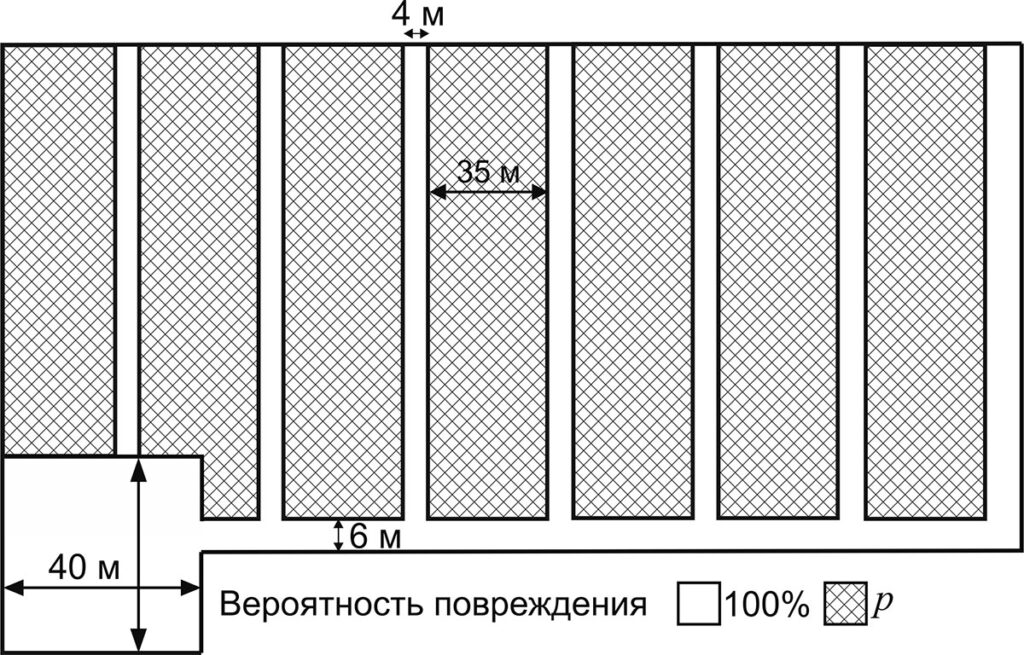
Рисунок 3. Моделируемая схема разработки лесосеки (с сохранением подроста методом узких пасек)
В связи с тем, что изменение экологических условий может оказывать существенное влияние на выживаемость ценопопуляций черники, при моделировании рубок учитывается увеличение влажности 20-сантиметрового слоя почвы на повреждённой территории на 50% (Пятецкий, Морозова, 1962; Пилепко, 2013). Значения влажности почвы возвращаются к исходным показателям линейно, в течение 12 лет в сосняках черничных и в течение 25 лет в ельниках черничных (Пятецкий, Морозова, 1962).
Моделирование реакции клонов черники на внешние воздействия. При моделировании реакции клонов черники на внешние воздействия парциальный куст считается погибшим при повреждении побеговой системы, либо при потере физиологической интеграции с другими парциальными кустами посредством корневищ. Восстановление клона начинается на следующем месячном шаге модели в случае повреждений в период вегетации, или в первый месяц следующего вегетационного периода при повреждениях в зимний период.
Поскольку целью данных имитационных экспериментов являлась оценка влияния нарушающих воздействий на динамику ценопопуляций в чистом виде, исключая воздействия, обусловленные динамикой древостоя, а также конкуренцией с другими видами растений травяно-кустарничкового яруса, экологические условия под пологом в ходе экспериментов оставались неизменными, а динамика других видов травяно-кустарничкового яруса не учитывалась. Экологические условия соответствовали средневозрастному сосняку, климатические условия соответствовали средним многолетним значениям для территории южного Подмосковья.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Реакция клонов черники на внешние воздействия. В ходе промежуточных наблюдений было выявлено, что отмирание парциальных кустов на площадках с имитацией повреждений происходит во временном интервале 2–8 недель с момента воздействия, вне зависимости от его характера. Новые парциальные кусты начали образовываться через 4 недели после нарушающих воздействий. На всех площадках, вне зависимости от характера повреждения, смена фенофаз происходила синхронно, генеративные особи плодоносили.
При наблюдении через 12 недель после закладки эксперимента были выявлены следующие закономерности. (1) При перерубании корневищ погибали парциальные образования, которые полностью потеряли связь с другими частями клона (выживаемость составила менее 3%). Выживаемость парциальных кустов, сохранивших связь хотя бы с ещё одним парциальным образованием, превышала 40%. (2) При перерубании корневищ и срезании побеговой системы происходило пробуждение от 1 до 3‑х спящих почек, расположенных на корневище на расстоянии 1–2 см от места повреждения (рис. 4А, 4Б). (3) При снятии лесной подстилки корневища, оказавшиеся на поверхности, принимали ортотропное положение, а парциальные образования, расположенные на них, образовывали с ранее функционирующими побегами единую надземную побеговую систему. При этом в ряде случаев у таких объединённых побегов на корневищах, расположенных в слое мха, происходило формирование новых корневищ на корневищах, принявших ортотропное положение (рис. 4В).

Рисунок 4. Пробудившиеся спящие почки (показаны стрелками) на месте срезанного парциального куста (А) и перерубленного корневища (Б); система побегов, образовавшаяся на корневище (показано стрелкой), принявшем ортотропное положение (В)
На всех корневищах, принявших ортотропное положение, наблюдалось образование новых корневищ из спящих почек, расположенных на расстоянии до 1 см от уровня почвы. Доля отмерших в зимний период парциальных кустов не превышала 5%. Соотношение погибших, выживших и новых парциальных кустов для каждого из типов воздействий, приведено на рисунке 5 и в таблице 2.
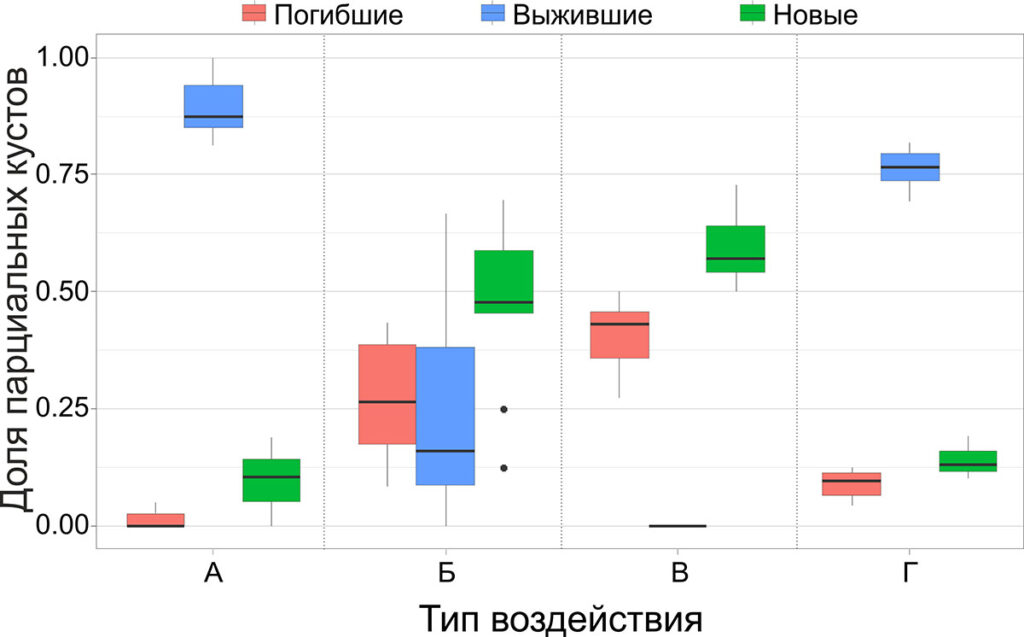
Рисунок 5. Соотношение погибших, выживших и новых парциальных кустов черники при различных типах воздействий (А – контроль, Б – перерубание корневищ, В – срезание побегов, Г – снятие подстилки, n=9)
Таблица 2. Соотношение погибших, выживших и новых парциальных кустов, шт. 0.25 м−2
| № ПП | До воздействия | После воздействия | ||
| Погибло | Выжило | Новых | ||
| Контроль | ||||
| 1 | 18 | 0 | 18 | 0 |
| 2 | 6 | 0 | 6 | 0 |
| 3 | 8 | 0 | 8 | 0 |
| 4 | 8 | 0 | 8 | 0 |
| 5 | 26 | 0 | 26 | 1 |
| 6 | 10 | 0 | 10 | 0 |
| 7 | 11 | 0 | 11 | 1 |
| 8 | 19 | 1 | 18 | 0 |
| 9 | 20 | 0 | 20 | 1 |
| Срезаны побеги | ||||
| 1 | 10 | 10 | 0 | 18 |
| 2 | 10 | 10 | 0 | 16 |
| 3 | 5 | 5 | 0 | 8 |
| 4 | 4 | 4 | 0 | 5 |
| 5 | 27 | 27 | 0 | 32 |
| 6 | 5 | 5 | 0 | 6 |
| 7 | 19 | 19 | 0 | 34 |
| 8 | 24 | 24 | 0 | 26 |
| 9 | 28 | 28 | 0 | 37 |
| Перерублены корневища | ||||
| 1 | 15 | 2 | 13 | 18 |
| 2 | 6 | 2 | 4 | 1 |
| 3 | 6 | 6 | 0 | 9 |
| 4 | 17 | 10 | 7 | 13 |
| 5 | 29 | 17 | 12 | 28 |
| 6 | 7 | 4 | 3 | 16 |
| 7 | 7 | 5 | 2 | 11 |
| 8 | 9 | 1 | 8 | 3 |
| 9 | 11 | 3 | 8 | 10 |
| Убрана лесная подстилка | ||||
| 1 | 42 | 9 | 33 (26*) | 8 |
| 2 | 17 | 0 | 17 (9*) | 0 |
| 3 | 20 | 0 | 20 (9*) | 2 |
| 4 | 12 | 0 | 12 (3*) | 0 |
| 5 | 44 | 2 | 42 (30*) | 2 |
| 6 | 14 | 1 | 13 (8*) | 3 |
| 7 | 16 | 0 | 15 (9*) | 1 |
| 8 | 24 | 0 | 24 (19*) | 3 |
| 9 | 23 | 1 | 22 (15*) | 0 |
* Число парциальных образований с учётом сформированной из ранее функционирующих побегов единой надземной побеговой системы
Более высокую выживаемость парциальных образований, сохранивших связь с другими частями клона, можно объяснить тем, что при физиологической интеграции парциальных образований ресурсы, получаемые неповреждённой частью клона, используются для восстановления и поддержания повреждённых парциальных образований. Такой эффект полностью согласуется с результатами, полученными ранее другими исследователями (Archer, Tieszen, 1980; Chapin, 1980a,b; Bryant et al., 1983).
Соотношение отмерших, выживших и новых парциальных кустов при снятии подстилки значимо не отличалось от контроля (p‑значение равно 0.437). При двух других типах воздействий отличия от контроля были значимыми (p-значение < 1⋅10−6), и была выявлена экспоненциальная сопряжённая зависимость количества новых парциальных образований от количества отмерших (уравнение 2).
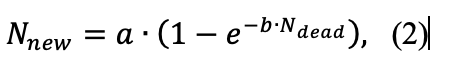
где Nnew – количество новых парциальных кустов, Ndead – количество отмерших парциальных кустов, a и b – эмпирические коэффициенты.
Количество ресурсов, которые неповреждённая часть клона способна потратить на восстановление своих повреждённых частей, ограничено. Как следствие, зависимость имеет нелинейный характер, при котором приращение функции убывает по мере увеличения количества отмерших парциальных кустов (рис. 6). При срезании побеговой системы партикуляция происходит с большей интенсивностью, и в данном случае функция выходит на насыщение позже. Это явление можно объяснить тем, что при уничтожении побеговой системы, дополнительные ресурсы не расходуются на восстановление тканей повреждённых парциальных образований, и, следовательно, появляется возможность сформировать большее количество новых парциальных кустов, что соотносится с литературными данными (Laine, 1988; Sonesson, Callaghan, 1991; Tolvanen, 1994).
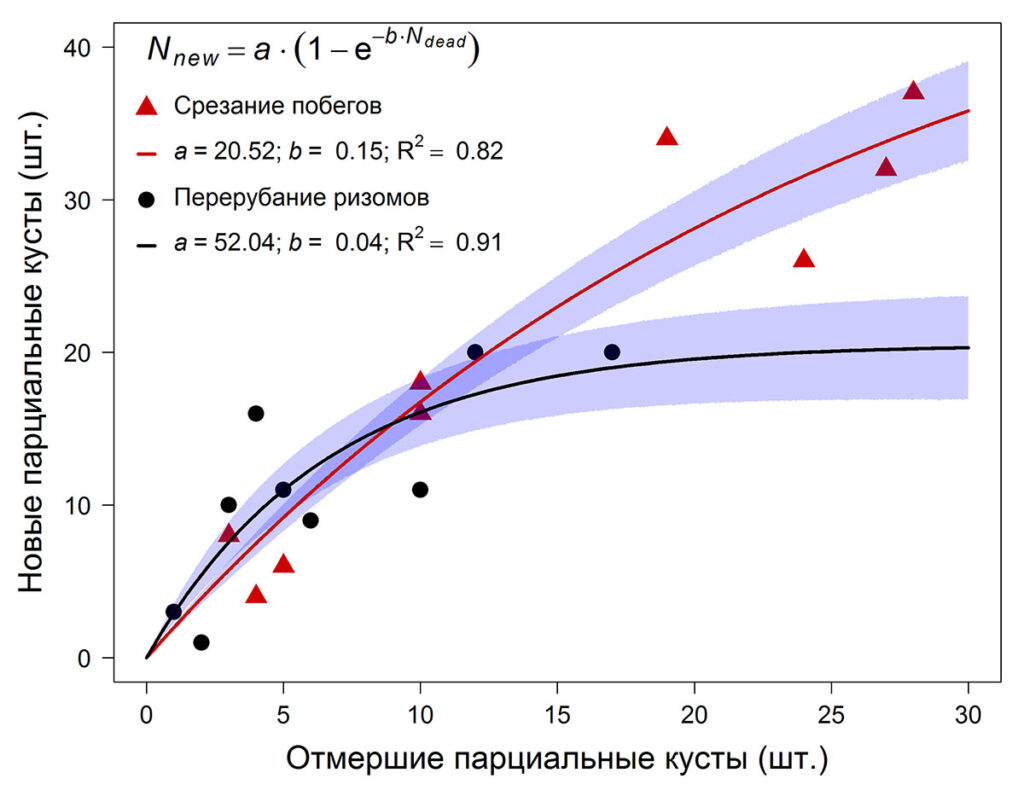
Рисунок 6. Зависимость количества новых парциальных кустов на площадке 0.25 м−2 от количества отмерших (причина отмирания указана на диаграмме). Заливкой показан 95% доверительный интервал.
Высокую выживаемость парциальных кустов черники при удалении лесной подстилки (более 75%) можно объяснить двумя факторами. Во-первых, при снятии подстилки была сохранена физиологическая интеграция корневищ на повреждённых участках с другими частями клона, следовательно, донорно-акцепторные взаимодействия позволили компенсировать нехватку минерального питания и воды парциальным кустам, лишённым возможности получать их напрямую. Во-вторых, количество осадков в летний период в 2016 году превышало климатическую норму в 1.46 раз (352 мм при норме 240 мм), что могло обеспечить необходимую влажность для корневищ, не прикрытых лесной подстилкой, препятствуя их засыханию.
Принятие корневищами ортотропного положения можно объяснить тем, что при снятии подстилки на периферии клона корневища переходят из диатропного (горизонтального) подземного в ортотропный ассимилирующий надземный побег – побег закрепления (Серебряков, Серебрякова, 1965), минуя период покоя (Маркаров, Головко, 1995). Потенциальная возможность такого явления у клонов черники показана в работе С. П. Масловой (2014). Данная способность позволяет парциальным кустам черники выживать при резких локальных изменениях экологических условий. Однако в литературных источниках приводятся данные о том, что, поскольку корневища черники расположены в слое мхов и лесной подстилки, большие площади нарушений, связанных с удалением подстилки, приводят к гибели корней и корневищ (Tolvanen, 1994; Hegland et al., 2005). В связи с этим применение полученных данных о реакции парциальных кустов черники на снятие лесной подстилки не представляется возможным при моделировании комплексных повреждений на большой территории.
Имитация отдельных видов воздействий. При помощи разработанной модели была проведена серия имитационных экспериментов по выявлению границ устойчивости ценопопуляций черники к внешним воздействиям, а также по определению скорости их восстановления после комплексных внешних воздействий. После внешних воздействий на этапе разрастания модифицируется схема расположения пробуждающихся спящих почек на корневище. Количество новых парциальных кустов, образующихся после повреждения части клона (Nnew), вычисляется в соответствии с уравнением 2, коэффициенты зависят от характера повреждений (рис. 6). При наличии повреждений и надземной, и подземной части значение Nnew вычисляется как минимальное между количеством новых парциальных кустов, образующихся при срезании побеговой системы и при перерубании корневищ. Таким образом, учитывается дополнительный расход ресурсов на восстановление повреждённых подземных частей клона. Ячейки, занятые только подземными органами черники в количестве Nnew, расположенные наиболее близко к повреждённой части клона, отмечаются как места пробуждения спящих почек. Плотность ценопопуляции оценивалась как количество парциальных кустов (шт.) на единицу площади (м2).
В первом имитационном эксперименте рассматривалась динамика ценопопуляции черники при отсутствии внешних воздействий и при повреждении различной интенсивности (1) побеговой системы (рис. 7) и (2) корневищ (рис. 8). Масштаб повреждений варьировал в диапазоне от 5 до 95% при разовых воздействиях и от 5 до 15% при систематических. Площадь моделируемой территории составляла 625 м2 (25 × 25 м), размер единичной ячейки был равен 1 см2. При инициализации модели территория заселялась семенами черники в количестве 50 шт. м−2. В ходе онтогенетического развития парциальных кустов учитывалась морфологическая и динамическая поливариантность онтогенеза. Моделирование проводилось на 100 лет (1200 шагов модели), разовые воздействия производились на 307‑м шаге (июль 25‑го года от начала моделирования), систематические воздействия – каждые 12 шагов, начиная с 307‑го. Во избежание эффектов, связанных с различным начальным расположением семян, затравочное число генератора псевдослучайных чисел было одинаковым во всех имитационных экспериментах.
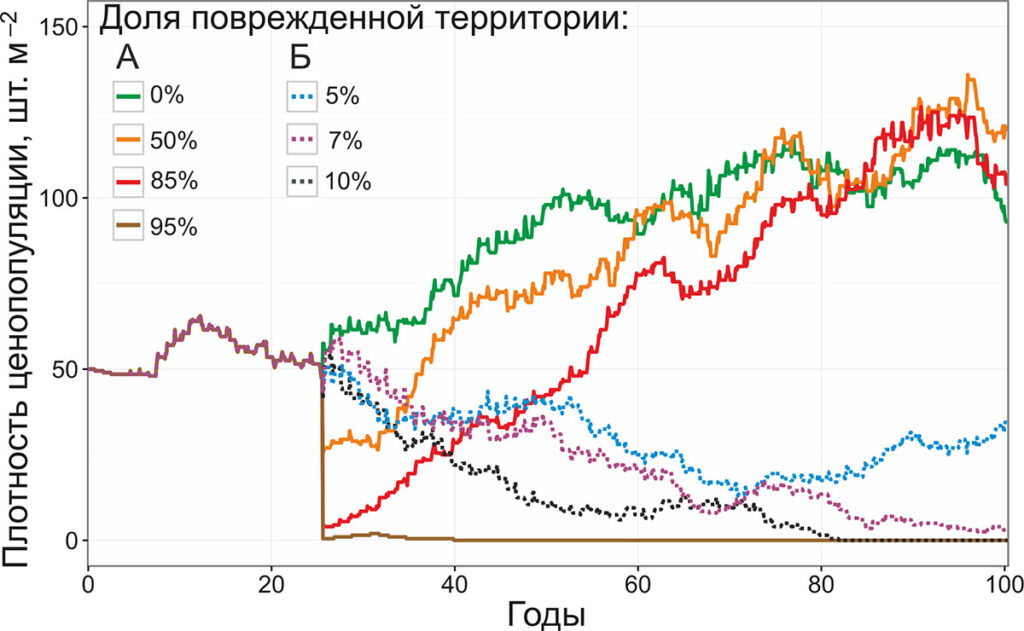
Рисунок 7. Динамика плотности ценопопуляции черники при повреждении побеговой системы (А – однократно, Б – ежегодно)
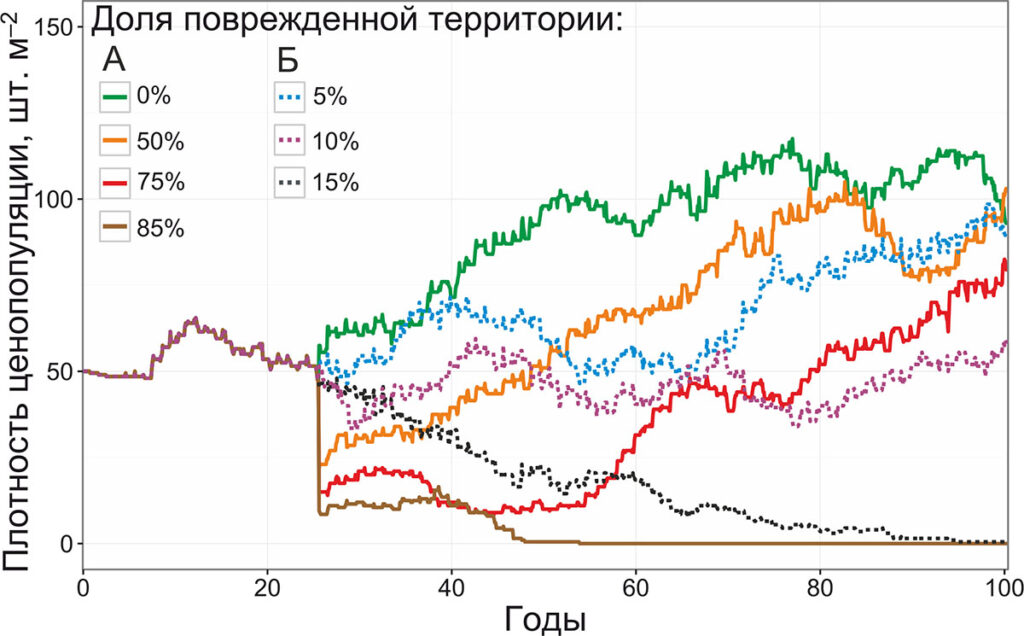
Рисунок 8. Динамика плотности ценопопуляции черники при повреждении корневищ (А – однократно, Б – ежегодно)
При моделировании развития ценопопуляции черники без внешних воздействий пиковая плотность ценопопуляции (116 шт. м−2) достигается к 75‑му году (900‑му шагу моделирования) и в дальнейшем удерживается на плато с незначительными колебаниями. При однократных повреждениях побеговой системы ценопопуляция черники показывает высокую устойчивость к внешним воздействиям, и погибает только при повреждении 95% парциальных кустов, тогда как воздействия меньшей интенсивности не приводят к исчезновению ценопопуляции. При повреждении 50% парциальных кустов восстановление плотности популяции до уровня контроля происходит в течение 35 лет, при повреждении 75% – в течение 55 лет. Плотность ценопопуляции черники при однократном воздействии с повреждением 10−15% ценопопуляции возрастает в связи с уменьшением внутривидовой конкуренции, связанной с пробуждением спящих почек вблизи материнского парциального образования. При систематических воздействиях наблюдается снижение устойчивости ценопопуляций к повреждению побегов. Так, гибель ценопопуляции черники происходит при повреждении 10% парциальных кустов ежегодно, плотность ценопопуляции в 4 раза ниже контроля наблюдается даже при ежегодном повреждении всего 5% парциальных кустов.
Устойчивость ценопопуляции черники к систематическим повреждениям подземных органов оказывается выше, чем к систематическим повреждениям побеговой системы. При повреждении подземных органов черники на 5% территории, плотность ценопопуляции достигает уровня контрольной ценопопуляции; исчезновение ценопопуляции происходит только при ежегодном повреждении 15% территории. Однако при разовых повреждениях подземных органов ценопопуляция черники гибнет при менее интенсивных воздействиях (повреждение 85% территории), чем при повреждении побеговой системы.
Высокую устойчивость ценопопуляции черники к повреждению побеговой системы парциальных кустов можно объяснить тем, что запаса питательных веществ, накопленных в корневищах, оказывается достаточно для пробуждения спящих почек. Однако при ежегодных воздействиях вновь образовавшиеся парциальные кусты не успевают перейти в виргинильное онтогенетическое состояние и начать вегетативное размножение, и в результате этого даже незначительные воздействия приводят к гибели ценопопуляции. При повреждении подземных органов, погибают только те парциальные кусты, которые теряют физиологическую интеграцию с другими, следовательно, при систематических повреждениях малой интенсивности доля погибших парциальных кустов оказывается ниже, чем при повреждении побеговой системы на той же территории. При увеличении доли территории, на которой повреждаются подземные органы, доля парциальных кустов, потерявших физиологическую интеграцию с соседями, увеличивается, и для восстановления ценопопуляции требуется большее количество ресурсов. В связи с этим устойчивость ценопопуляции к единовременным повреждениям подземных органов высокой интенсивности оказывается ниже, чем к повреждению побеговой системы.
Имитация низовых лесных пожаров. Во втором имитационном эксперименте рассматривалась динамика ценопопуляции черники в терминах плотности популяции при пожарах различной способности к распространению (от 0.01 до 0.15 с шагом 0.01). Частота пожаров составляла 1 раз в 5 лет, начиная с 305‑го шага (май 25‑го года от начала моделирования). Значение параметра n (ширина зоны устойчивых к возгоранию ячеек, при котором распространение пожара прекращалось в данном направлении), было задано равным 200 (2 метра). Многолетняя среднемесячная влажность лесной подстилки в мае составляла 10.3 ± 1.2 об. %. Моделируемая территория являлась пространственно-неоднородной по почвенно-климатическим условиям (значения температуры и влажности в разных ячейках варьировали в пределах 10% от базовых значений). Климатические условия соответствовали территории южного Подмосковья (Булыгина и др., 2019; Шерстюков, 2019). Площадь моделируемой территории составляла 10000 м2 (100 × 100 м). При инициализации модели территория заселялась семенами черники в количестве 50 шт. м−2. Для остальных параметров были заданы те же значения, что и в первом эксперименте. Результаты эксперимента приведены на рисунке 9.
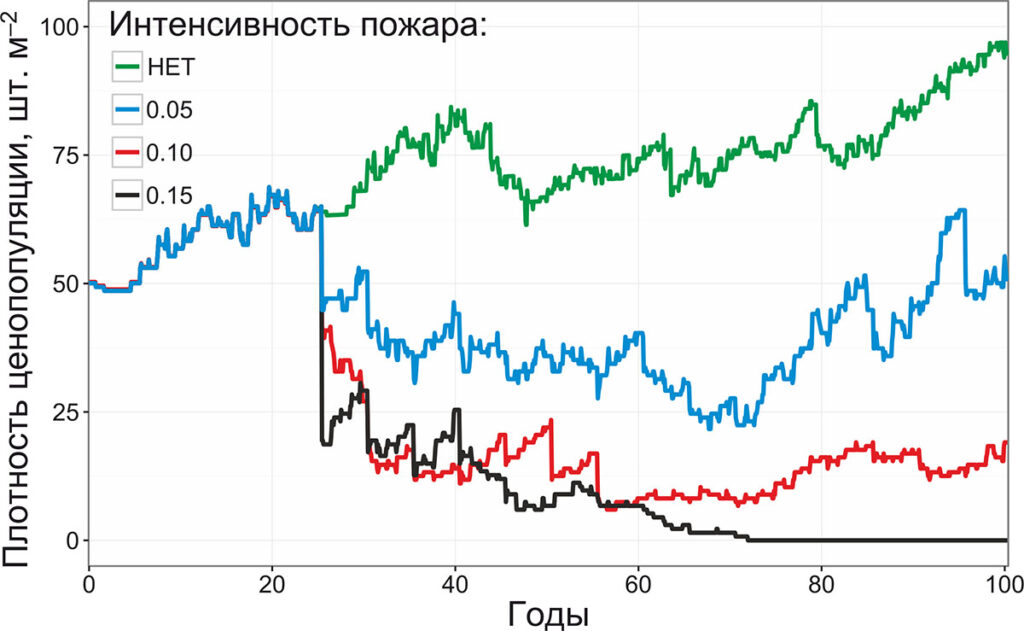
Рисунок 9. Динамика плотности ценопопуляции черники при низовых лесных пожарах различной способности к распространению
Основным отличием данного эксперимента от случайного повреждения побеговой системы с заданной вероятностью являлось неслучайное расположение повреждаемой территории. Парциальные кусты черники, расположенные в наиболее влажных местах, не повреждались. В связи с этим, при моделировании пожаров со способностью к распространению 0.05 и 0.1 (повреждающих ячейки с объёмной влажностью подстилки ≤ 5% и 10% соответственно), плотность ценопопуляции черники выходила на плато с периодическими колебаниями. Ни растущего, ни снижающегося тренда при этом не наблюдалось. Гибель ценопопуляции черники при способности пожара к распространению 0.15 происходит к 73‑му году (876‑му шагу моделирования) постепенно, резкие пики снижения плотности ценопопуляции наблюдаются только на 25‑м, 30‑м, 35‑м и 40‑м году. Постепенная гибель ценопопуляции может говорить о том, что причиной гибели явилось не единовременное сгорание побеговых систем всех парциальных кустов, а малая площадь участков территории, не повреждаемой пожарами, которой оказывается недостаточно для самоподдержания ценопопуляции.
Имитация выборочных рубок. В третьем имитационном эксперименте рассматривалась динамика ценопопуляции черники в терминах плотности популяции при выборочных рубках в сосняках методом узких пасек в соответствии со схемой, приведённой на рисунке 6, с применением техники (1) на шинах низкого давления и (2) на гусеничном шасси. Рубки имитировались на 25‑м году моделирования в зимний (300‑й шаг модели, толщина снежного покрова – 0.13 м, мощность лесной подстилки 0.05 м) и летний (307‑й шаг модели, мощность лесной подстилки 0.05 м) период. Плотность подстилки и снежного покрова составляли 151 и 243 кг м−3, соответственно. Площадь моделируемой территории составляла 100000 м2 (200 × 500 м). При инициализации модели территория заселялась семенами черники в количестве 50 шт. м−2. Для остальных параметров были заданы те же значения, что и во втором эксперименте. Результаты эксперимента приведены на рисунке 10.
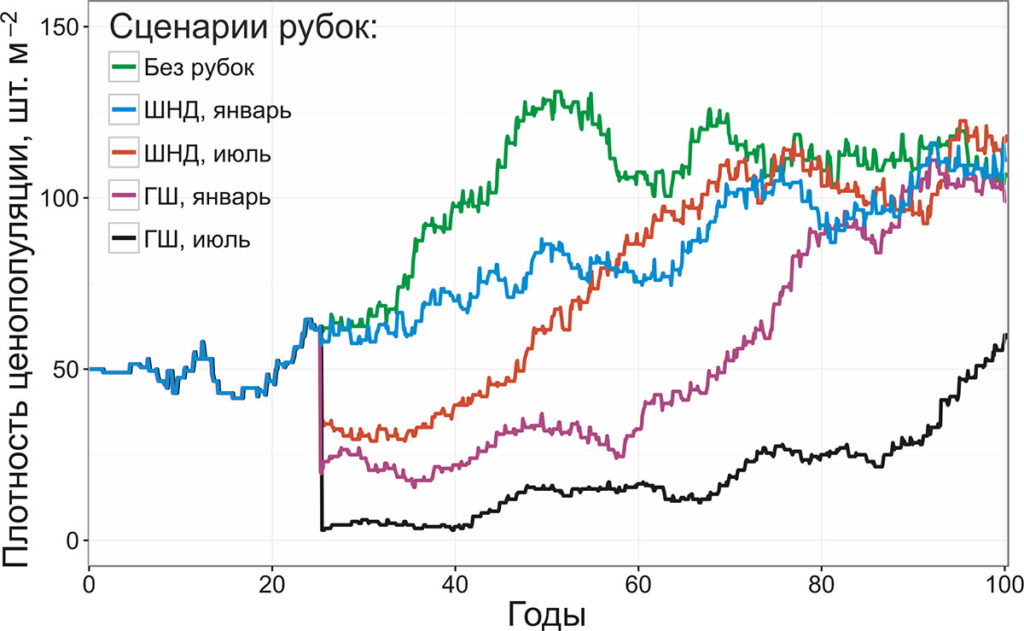
Рисунок 10. Динамика плотности ценопопуляции черники при различных особенностях рубок
В ходе имитационного эксперимента было установлено, что ни один из рассмотренных видов рубок не приводил к гибели ценопопуляции черники. В случае рубки с применением техники на шинах низкого давления в зимний период резкого снижения плотности ценопопуляции не наблюдалось, однако по сравнению с контрольной ценопопуляцией рост плотности происходил медленнее. При моделировании рубок с применением техники на шинах низкого давления в летний период, а также техники на гусеничном шасси в зимний период наблюдается единовременное резкое снижение плотности ценопопуляции с последующим восстановлением до значений контрольной ценопопуляции в течение 44–46 лет. Применение техники на гусеничном шасси в летний период приводит к повреждению порядка 80% моделируемой территории, восстановление ценопопуляции черники происходит медленнее, чем при других вариантах рубок, плотность ценопопуляции оказывается ниже контрольной в 1.9 раз через 75 лет после воздействия. Увеличившаяся влажность лесной подстилки после рубок не оказала существенного воздействия на восстановление ценопопуляции черники, т.к. оставалась в пределах диапазона толерантности (менее 64%).
ЗАКЛЮЧЕНИЕ
Проведённое исследование с использованием модели CAMPUS‑S позволило оценить устойчивость ценопопуляций черники к различным типам нарушающих воздействий при низовых пожарах и выборочных рубках, таким как повреждение побеговой системы и корневищ, удаление подстилки. Ценопопуляции черники демонстрируют высокую устойчивость к разовым повреждениям побеговой системы, восстанавливаясь даже при уничтожении до 85% парциальных кустов. Однако систематические воздействия приводят к постепенному снижению плотности популяции, а при повреждении более 10% территории – к её гибели. Повреждение подземных органов оказывается менее критично при систематических воздействиях малой интенсивности, но разовые масштабные нарушения, затрагивающие более 85% территории, приводит к гибели ценопопуляции. Низовые пожары со средней способностью к распространению (до 10% повреждения территории) не приводят к гибели ценопопуляции при условии частичного сохранения парциальных образований черники в понижениях микрорельефа, но при увеличении способности к распространению до 15% восстановление становится невозможным. Выборочные рубки, даже с применением тяжёлой техники, не вызывают полной гибели ценопопуляции черники, но замедляют её восстановление. Ожидаемо, наиболее значительное снижение плотности наблюдается при рубках в летний период с использованием гусеничной техники.
Ключевым фактором выживания черники является физиологическая интеграция клона, позволяющая перераспределять ресурсы между повреждёнными и неповреждёнными частями, а активация спящих почек на корневищах и формирование новых парциальных кустов обеспечивают восстановление популяции после повреждений.
Применённая в данном исследовании модель имеет ряд ограничений. Во-первых, высокое пространственное разрешение ограничивает её применение для больших территорий из-за вычислительной сложности. Во-вторых, распределение повреждений при пожарах и рубках предполагает идеализированные условия, что может не полностью соответствовать реальной неоднородности лесных экосистем, а функции, используемые при имитации повреждений вследствие пожаров и рубок, требуют дополнительной параметризации и валидации. В-третьих, при проведении имитационных экспериментов древостой, другие виды растений травяно-кустарничкового яруса, и, следовательно, сукцессионная динамика, не учитывались, поскольку могли оказать существенное влияние на динамику ценопопуляций черники и являлись бы дополнительными факторами, определяющими восстановление после внешних воздействий. В то же время, данные факторы могут играть ключевую роль в динамике ценопопуляций черники.
Несмотря на упомянутые выше ограничения, при параметризации других видов доминантов ТКЯ и учёте имеющихся ограничений, модель CAMPUS‑S, дополненная модулем внешних воздействий, может использоваться для (1) прогнозирования динамики ценопопуляций растений ТКЯ при различных сценариях антропогенных и природных воздействий, (2) оценки устойчивости лесных экосистем к нарушениям, (3) разработки рекомендаций по их сохранению и (4) планирования лесохозяйственных мероприятий, таких как рубки и противопожарные меры, с учётом минимизации ущерба для ТКЯ.
ФИНАНСИРОВАНИЕ
Полевые исследования выполнены в рамках государственного задания ФИЦ ПНЦБИ РАН (рег. № 122040500037-6), моделирование – в рамках государственного задания ЦЭПЛ РАН «Биоразнообразие и экосистемные функции лесов» (рег. № 124013000750-1).
СПИСОК ЛИТЕРАТУРЫ
Бобкова К. С. Биологическая продуктивность хвойных лесов европейского Северо-Востока. Л.: Наука, 1987. 156 с.
Бойчук А. П., Дроздов А. В. Оценка работы систем машин на добровольно-выборочных рубках // Эколого-географические проблемы сохранения и восстановления лесов Севера. Архангельск: АИЛиЛХ, 1991. С. 326–328.
Булыгина О. Н., Разуваев В. Н., Трофименко Л. Т., Швец Н. В. Описание массива данных среднемесячной температуры воздуха на станциях России: свидетельство о гос. регистрации базы данных № 2014621485. – URL: http://meteo.ru/data/156-temperature (дата обращения: 12.12.2019).
Валова З. Г. Рубки ухода и урожай черники // Лесохозяйственная наука и практика. Минск: Урожай, 1973. С. 46–57.
Колычева А. А., Чумаченко С. И. Потенциал урожая черники, брусники, малины с учетом особенностей участка и сценария ведения лесного хозяйства // Научные основы устойчивого управления лесами: материалы Всероссийской научной конференции с международным участием. М.: ЦЭПЛ РАН, 2022. С. 167–170.
Комаров А. С. Простые структуры растительного покрова, устойчивые к внешним нарушениям // Взаимодействующие марковские процессы и их применение к математическому моделированию биологических систем. Пущино: ПНЦ РАН, 1982. С. 136–143.
Комаров А. С., Паленова М. М. Моделирование взаимодействующих популяций вегетативно-подвижных трав // Бюллетень МОИП. Отд. биол. 2001. Т. 106. № 5. С. 34–41.
Комарова Т. А. Изменение синузиальной структуры кустарничково-травяного яруса в ходе послепожарных сукцессий // Ботанический журнал. 1993. Т. 78. № 6. С. 86–95.
Мариничев Е. А., Мариничева Т. В. Естественное возобновление на лесосеках сплошных рубок, разработанных методом узких лент // Вестник Нижегородской государственной сельскохозяйственной академии. 2016. № 1. С. 19–22.
Маркаров А. М., Головко Т. К. Ростовая ориентация подземных побегов многолетних травянистых растений. 3. Морфофизиология подземных побегов и развитие сарментов // Физиология растений. 1995. Т. 42. № 5. С. 709–713.
Маслова С. П. Экофизиология подземного метамерного комплекса длиннокорневищных растений: Дисс. … д-ра биол. наук (спец. 03.01.05 – «Физиология и биохимия растений», 03.02.08 – «Экология» (в биологии)). СПб., 2014. 233 с.
Михайлова Н. В., Михайлов А. В., Богданова Н. Е., Комаров А. С., Жукова Л. А. Имитационная модель инвазионной динамики популяций неморальных видов трав на неоднородной территории // Бюллетень МОИП. Отд. биол. 2008. Т. 113. № 5. С. 68–75.
Пилепко Е. Н. Экологическое обследование вырубок на Онежском полуострове // Materiały z IX Międzynarodowej naukowo-praktycznej konferencji «Kluczowe aspekty działalności naukowej – 2013». Polska, 2013. P. 40–44.
Пятецкий Г. Е., Морозова Д. М. Изменение физических и химических свойств лесных почв южной Карелии в связи с вырубками леса // Лесные почвы Карелии и изменение их под влиянием лесохозяйственных мероприятий. Вып. 34. Петрозаводск, 1962. С. 71–92.
Рожин Л. П., Григорьев М. Н. Лесоводственная оценка машины ВТМ‑4 // Лесная промышленность. 1972. № 6. С. 17–18.
Серебряков И. Г., Серебрякова Т. И. О двух типах формирования корневищ у травянистых многолетников // Бюллетень МОИП. Отд. биол. 1965. Т. 70. № 1. С. 67–81.
Шерстюков А. Б. Описание массива суточных данных о температуре почвы на глубинах до 320 см по метеорологическим станциям Российской Федерации (версия 2). – URL: http://meteo.ru/data/164-soil-temperature (дата обращения: 12.12.2019).
Archer S., Tieszen L. L. Growth and physiological responses of tundra plants to defoliation // Arctic and Alpine Research. 1980. Vol. 12. No. 4. P. 531–552. DOI: 10.1080/00040851.1980.12004211
Atlegrim O. Interactions between the bilberry (Vaccinium myrtillus) and a guild of insect larvae in a boreal coniferous forest: PhD thesis. Umeå: Swedish University of Agricultural Sciences, 1991. 26 р.
Bryant J. P., Chapin F. S., Klein D. R. Carbon nutrient balance of boreal plants in relation to vertebrate herbivory // Oikos. 1983. Vol. 40. No. 3. P. 357–368. DOI: 10.2307/3544308
Cargill S. M., Jeffries R. L. Nutrient limitation of primary production in a sub-arctic salt marsh // Journal of Applied Ecology. 1984. Vol. 21. No. 2. P. 657–668. DOI: 10.2307/2403436
Chapin F. S. Effect of clipping upon nutrient status and forage value of tundra plants in Arctic Alaska // Proceedings of the 2nd International Reindeer/Caribou Symposium (17–21 September 1979). Røros, 1980b. P. 19–25.
Chapin F. S. Nutrient allocation and responses to defoliation in tundra plants // Arctic and Alpine Research. 1980a. Vol. 12. No. 4. P. 553–563. DOI: 10.2307/1550500
Chertov O., Komarov A., Shaw C., Bykhovets S., Frolov P., Shanin V., Grabarnik P., Priputina I., Zubkova E., Shashkov M. Romul_Hum – A model of soil organic matter formation coupling with soil biota activity. II. Parameterisation of the soil food web biota activity // Ecological Modelling. 2017a. Vol. 345. P. 125–139. DOI: 10.1016/j.ecolmodel.2016.10.024
Chertov O., Shaw C., Shashkov M., Komarov A., Bykhovets S., Shanin V., Grabarnik P., Frolov P., Kalinina O., Priputina I., Zubkova E. Romul_Hum model of soil organic matter formation coupled with soil biota activity. III. Parameterisation of earthworm activity // Ecological Modelling. 2017b. Vol. 345. P. 140–149. DOI: 10.1016/j.ecolmodel.2016.06.013
Dahlgren J., Oksanen L., Sjödin M., Olofsson J. Interactions between gray-sided voles (Clethrionomys rufucanus) and bilberry (Vaccinium myrtillus), their main winter food plant // Oecologia. 2007. Vol. 152. P. 525–532. DOI: 10.1007/s00442-007-0664-8
Emanuelsson U. Ecological effects of grazing and trampling on mountain vegetation in Northern Sweden: PhD thesis. Lund: University of Lund, 1984.
Ericson L. The influence of voles and lemmings on the vegetation in a coniferous forest during a 4-year period in northern Sweden // Wahlenbergia Scripta Botanica Umensia. 1977. Vol. 4. P. 11–14.
Flower‑Ellis J. G. K. Age structure and dynamics in stands of bilberry (Vaccinium myrtillus L.): PhD thesis. Stockholm: Royal College of Forestry, 1971.
Frolov P. V., Shanin V. N., Zubkova E. V., Bykhovets S. S., Grabarnik P. Y. CAMPUS-S – The model of ground layer vegetation populations in forest ecosystems and their contribution to the dynamics of carbon and nitrogen. I. Problem formulation and description of the model // Ecological Modelling. 2020a. Vol. 431. ID 109184. DOI: 10.1016/j.ecolmodel.2020.109184
Frolov P. V., Zubkova E. V., Shanin V. N., Bykhovets S. S., Mäkipää R., Salemaa M. CAMPUS-S – The model of ground layer vegetation populations in forest ecosystems and their contribution to the dynamics of carbon and nitrogen. II. Parameterization, validation and simulation experiments // Ecological Modelling. 2020b. Vol. 431. ID 109183. DOI: 10.1016/j.ecolmodel.2020.109183
Haag R. W. Nutrient limitations to plant production in two tundra communities // Canadian Journal of Botany. 1974. Vol. 52. No. 1. P. 103–116. DOI: 10.1139/b74-014
Havas P. The water economy of the bilberry (Vaccinium myrtillus) under winter conditions // Report from Kevo Subarctic Research Station. 1971. Vol. 8. P. 41–52.
Hegland S. J., Rydgren K., Seldal T. The response of Vaccinium myrtillus to variations in grazing intensity in a Scandinavian pine forest on the island of Svanøy // Canadian Journal of Botany. 2005. Vol. 83. No. 12. P. 1638–1644. DOI: 10.1139/b05-132
Komarov A., Chertov O., Bykhovets S., Shaw C., Nadporozhskaya M., Frolov P., Shashkov M., Shanin V., Grabarnik P., Priputina I., Zubkova E. Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing // Ecological Modelling. 2017. Vol. 345. P. 113–124. DOI: 10.1016/j.ecolmodel.2016.08.007
Laine K. Long-term variations in plant quality in relation to cyclic microtine rodents at Kilpisjärvi, Finnish Lapland: PhD thesis. Oulu: University of Oulu, 1988.
Matsumoto M., Nishimura T. Mersenne twister: a 623-dimensionally equidistributed uniform pseudorandom number generator // ACM Transactions on Modeling and Computer Simulation. 1998. Vol. 8. No. 1. P. 3–30. DOI: 10.1145/272991.272995
Mattson W. J. Herbivory in relation to plant nitrogen content // Annual Review of Ecology and Systematics. 1980. Vol. 11. No. 1. P. 119–161. DOI: 10.1146/annurev.es.11.110180.001003
Morén A. S., Lindroth A. CO2 exchange at the floor of a boreal forest // Agricultural and Forest Meteorology. 2000. Vol. 101. No. 1. P. 1–14. DOI: 10.1016/S0168-1923(99)00160-4
Oborny B., Englert P. Plant growth and foraging for a patchy resource: a credit model // Ecological Modelling. 2012. Vol. 234. P. 20–30. DOI: 10.1016/j.ecolmodel.2011.12.017
Pakonen T., Saari E., Laine K., Havas P., Lathdesmgki P. How do seasonal changes in carbohydrate concentrations in tissues of the bilberry (Vaccinium myrtillus L.) reflect carbon resource allocation patterns? // Acta Oecologica. 1991. Vol. 12. P. 249–259.
R Core Team. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing, 2022. – URL: https://www.R-project.org/ (accessed on: 12.12.2022).
Sjörs H. Vaccinium myrtillus, a plant portrait // Svensk Botanisk Tidskrift. 1989. Vol. 83. P. 411–428.
Sonesson M., Callaghan T. V. Strategies of survival in plants of the Fennoscandian tundra // Arctic. 1991. Vol. 44. No. 2. P. 95–105. DOI: 10.14430/arctic1525
Stephen D., Adruce S. A. Z. Cochran’s Q with pairwise McNemar for dichotomous multiple responses data: a practical approach // International Journal of Engineering and Technology. 2018. Vol. 7. No. 3. P. 4–6. DOI: 10.14419/ijet.v7i3.18.16662
Tolvanen A. Differences in recovery between a deciduous and an evergreen ericaceous clonal dwarf shrub after simulated aboveground herbivory and belowground damage // Canadian Journal of Botany. 1994. Vol. 72. No. 6. P. 853–859. DOI: 10.1139/b94-110
Рецензент: д. б. н., доцент, зав. кафедрой Чумаченко С. И.





