- DOI: 10.31509/2658-607x-202583-172
- УДК 630*232.3
Восстановление кедрово-широколиственного леса после однократных рубок 1960-х годов в южной части Сихотэ-Алиня
![]()
© 2025 О. Н. Ухваткина*, А. М. Омелько, В. Е. Захарова, А. А. Жмеренецкий,
Г. А. Гладкова, Л. А. Сибирина, А. В. Куприн
Федеральный научный центр Биоразнообразия наземной биоты Восточной Азии ДВО РАН
Россия, 690022, г. Владивосток, пр. 100 лет Владивостоку, д. 159
*E-mail: ukhvatkina@biosoil.ru
Поступила в редакцию 25.07.2025
После рецензирования: 09.08.2025
Принята к печати: 15.08.2025
Актуальность и цель. Кедрово-широколиственные леса южного Сихотэ-Алиня являются уникальными лесными экосистемами с высокой биологической ценностью и сложной структурой. В условиях продолжающегося хозяйственного освоения и деградации коренных лесов оценка долгосрочных последствий прошлых рубок приобретает особую значимость. Целью данного исследования является анализ состояния древостоев спустя 60–70 лет после однократных приисковых рубок, проведённых в 1960-х гг., и сравнение их с сохранившимися старовозрастными древостоями. Материал и методы. Материалом для исследования послужили данные, собранные на территории Верхнеуссурийского биогеоценотического стационара, где было заложено 346 временных пробных площадей. Для анализа отобраны 127 пробных площадей с типом леса К4 (разнокустарниковый кедровник с берёзой жёлтой), из них 58 находились в послерубочном древостое и 69 – в старовозрастном. Измерялись диаметр и высота деревьев, рассчитывалась площадь поперечного сечения и объём ствола деревьев. Статистический анализ выполнен с использованием непараметрических статистических методов в среде Python. Результаты. Полученные результаты показывают, что даже через 60–70 лет после рубок сохраняются существенные отличия от старовозрастных древостоев: ниже запас древесины, меньше среднее значение диаметра деревьев и отсутствуют крупные деревья. В видовом составе доминируют быстрорастущие лиственные виды, тогда как доля кедра корейского значительно снижена. Заключение. Восстановление исходной структуры кедрово-широколиственных лесов происходит крайне медленно и остаётся незавершённым даже спустя несколько десятилетий после рубок. В связи с этим необходимо пересмотреть план лесохозяйственных мероприятий в целях сохранения и восстановления этих экосистем.
Ключевые слова: Кедрово-широколиственные леса, однократные рубки, восстановление древостоя, южный Сихотэ-Алинь, видовой состав
Лесные экосистемы выполняют ключевую роль в устойчивости ландшафтов, сохранении биоразнообразия и обеспечении человека природными ресурсами. Поэтому в условиях масштабной антропогенной трансформации, вызванной интенсивными рубками и пожарами, вопросы рационального лесопользования остаются особенно актуальными, в том числе в Дальневосточном регионе России (Данилин, 2004; Ковалев, 2004). Одной из наиболее уязвимых и одновременно ценных лесных формаций являются кедрово-широколиственные леса. Эти леса обеспечивают не только высокое видовое разнообразие, но и служат ключевым элементом экосистемной устойчивости, формируя кормовую базу для промысловых животных, включая копытных, чья численность непосредственно влияет на состояние популяции амурского тигра – редчайшего хищника, находящегося под угрозой исчезновения (Куренцова, 1968).
Кедрово-широколиственные леса с сосной корейской (кедром, Pinus koraiensis Siebold et Zucc.) характеризуются сложной возрастной структурой и мозаичным распределением доминантов (Колесников, 1956). Однако вследствие исторически высоких нагрузок на лесные ресурсы, особенно в период массовых заготовок древесины в середине XX в., значительные площади коренных кедровников были трансформированы в производные леса (Соловьев, 1948; Розенберг, 1975). Несмотря на попытки регламентировать хозяйственную деятельность, промышленные рубки в XX в. привели к деградации местообитаний, снижению полноты древостоев и сокращению в них участия кедра корейского (Гуков, 1989).
Процессы восстановления после таких рубок, как показывают классические (Соловьев, 1948) и современные исследования (Комарова и др., 2022), носят длительный и неоднозначный характер. Начальные стадии лесовосстановления обычно характеризуются преобладанием быстрорастущих лиственных видов — в первую очередь берёзы жёлтой (Betula costata Trautv.) и тополя дрожащего (Populus tremula L.), тогда как участие хвойных видов, особенно кедра, остаётся низким. Естественное восстановление кедра осложняется рядом факторов: низкой семенной продуктивностью, эпизодическим плодоношением, а также поеданием семян грызунами (Усенко, 1984; Кудинов, 2007). Без специальных лесоводственных мероприятий возврат к исходной структуре кедрово-широколиственного леса в обозримой перспективе представляется маловероятным (Лебедев, 2003).
На этом фоне возрастает значимость оценки состояния древостоев, пройденных однократными промышленными рубками более полувека назад. Такие участки при отсутствии пожаров и иных нарушений представляют собой уникальную возможность проследить долгосрочные сукцессионные процессы и оценить потенциал естественного восстановления коренных лесов. Особенно важно провести их сравнение с сохранившимися старовозрастными древостоями, не подвергавшимися хозяйственному воздействию (Громцев, 1999). В то же время найти такие участки представляется крайне непростой задачей, поскольку большая часть площади лесного фонда Приморского края за последние 70-80 лет была пройдена рубками более четырех-пяти раз (Ковалев, Качанова 2023), что вносит значительные корректировки в процессы лесовосстановления.
Несмотря на это на территории Верхнеуссурийского биогеоценотического стационара – уникального научного полигона в южной части Сихотэ-Алиня, где представлены как участки коренных кедрово-широколиственных и елово-широколиственных лесов, так и участки, однократно пройденные рубками в 1960-х годах (Архивы ДВО РАН, отчеты ВУС 1964-1965 гг.; Сибирина и др., 2022). Это позволяет в сопоставимых условиях оценивать структуру древостоев, участие видов-доминантов, наличие подроста и особенности видового состава на разных этапах восстановления. Цель исследования – оценить успешность восстановления древостоев на участках, пройденных однократными рубками в конце 1960-х годов.
МАТЕРИАЛ И МЕТОДЫ
Территория исследования. Верхнеуссурийский стационар ФНЦ Биоразнообразия ДВО РАН расположен в бассейне реки Правая Соколовка (Приморский край, 44°02′ N, 134°12′ E, рис. 1.). Стационар охватывает горные участки на высотах от 460 до 1060 м над ур. м. и характеризуется высокой лесистостью, мозаичным рельефом и разнообразием фитоценозов. Преобладающими типами лесной растительности являются кедрово-широколиственные, смешанные широколиственные и елово-пихтовые леса (Яковлева, 2004). Климат умеренно муссонный, с годовой суммой осадков около 830 мм и выраженным летним максимумом. Среднегодовая температура воздуха составляет 0.9 °С (Кожевникова, 2009). Продолжительность вегетационного периода составляет 120-140 дней.
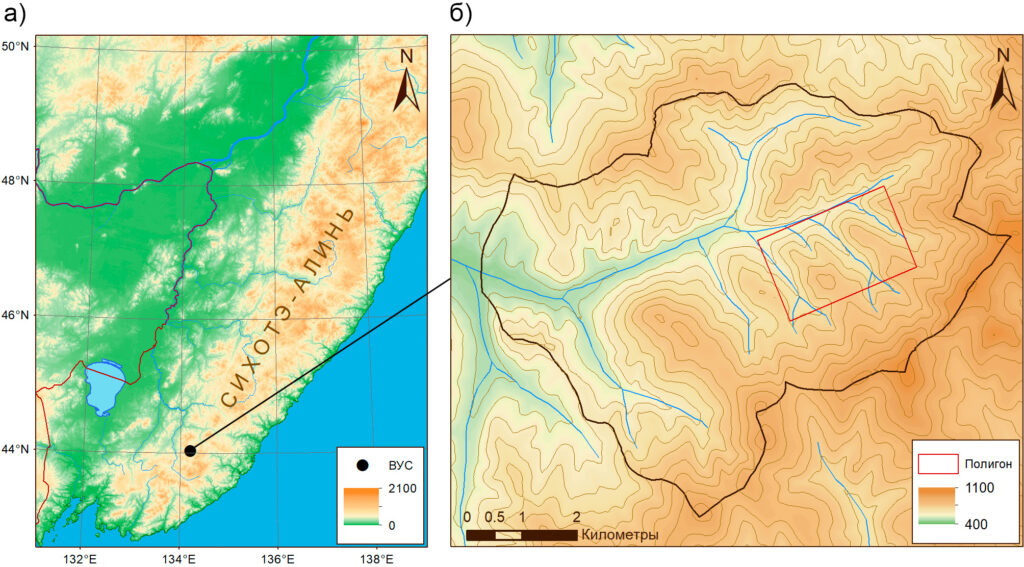
Рисунок 1. Территория исследования: а – расположение Верхнеуссурийского стационара ФНЦ Биоразнообразия ДВО РАН в пределах Сихотэ-Алиня, б – схема территории стационара и расположение тестового полигона
Рубки в регионе и на территории стационара. На большей части территории Приморского края интенсивные заготовки древесины велись начиная с середины XX в., включая массовое освоение труднодоступных горных лесов в 1950–1970-х годах. В пределах Верхнеуссурийского стационара крупномасштабные лесозаготовки проводились в основном в 1960-х годах. Для анализа лесовосстановления после рубок в лесах с преобладанием кедра корейского были выделены участки, пройденные в 1965–1969 гг. выборочными рубками приискового характера, направленными на заготовку лучших стволов кедра корейского, а также сплошные рубки в темнохвойных лесах с преобладанием ели аянской. В то же время, в отдалённых и труднодоступных местах стационара сохранились участки древостоев, не подвергавшиеся вырубкам и пожарам. Они были использованы как эталонные участки.
Сбор данных. На территории стационара был заложен тестовый полигон площадью 400 га (размеры 2.5 × 1.6 км), охватывающий мозаику лесов разного типа. С помощью алгоритмической кластеризации данных дистанционного зондирования полигон был разделён на 346 элементарных участков площадью около 1 га. На каждом из них закладывалась круговая временная пробная площадь радиусом 11.3 м (площадь 0.04 га).
В пределах временной пробной площади проводилось комплексное описание древостоя. Для всех деревьев определялся вид, состояние (живое: нормальное или фаутное; мёртвое: сухостой), диаметр на высоте груди (DBH). Для части деревьев дополнительно измерялась высота. В дальнейшем по сочетанию диаметра и разряда высот с использованием стандартных таблиц объёмов стволов рассчитывался объём ствола каждого отдельного дерева.
В ходе работ на пробных площадях проводился поиск следов рубок (пни, следы дорог или волоков и т.д.). По их наличию или отсутствию все участки были разделены на две категории: 1) старовозрастные – древостой без следов рубок или пожаров, 2) послерубочные – древостой пройден рубками в 1960-х годах. По данным лесоустройства, которое на территории стационара было выполнено по I разряду, участки были отнесены к разным типам леса. Для статистического анализа был выбран тип леса К4 (разнокустарниковый кедровник с берёзой жёлтой), поскольку он, с одной стороны, оказался наиболее распространённым (127 из 346 участков), и, с другой стороны, для данного типа было сравнимое число как старовозрастных, так и послерубочных участков (69 и 58 соответственно).
Статистический анализ. Анализ проводился по двум «уровням»: 1) общие характеристики древостоя по категориям участков (старовозрастные и послерубочные), 2) сравнение показателей по отдельным видам деревьев. Настоящее исследование было сосредоточено на анализе структуры древостоя, поскольку его характеристики наиболее репрезентативны для оценки различий между категориями участков, поэтому анализ подроста не проводился. Также, учитывая небольшой размер временных пробных площадей (0.04 га), ограничивающий полноту оценки структуры и видового состава при высоком видовом разнообразии, характерном для кедрово-широколиственных лесов, для анализа характеристик древостоев все временные пробные площади в пределах каждой категории (старовозрастные / послерубочные) были агрегированы. В первом случае суммарная площадь составила 2.76 га, во втором – 2.32 га.
На первом этапе был составлен перечень древесных видов, встречающихся на старовозрастных и послерубочных участках, для последующего сравнения. Затем, отдельно для каждой категории участков, были рассчитаны следующие показатели: число деревьев, сумма площадей поперечного сечения (basal area — BA), запас древостоя, средние и медианные значения диаметра, высоты и объёма ствола. Все значения нормированы на 1 га. Распределения признаков оценивались с использованием гистограмм и функций накопленного распределения (CDF). Для статистической проверки различий между группами применялись непараметрические методы: тест Манна–Уитни — для сравнения медианных значений, тест Колмогорова–Смирнова — для анализа различий в форме распределений.
Дополнительно проводился анализ на уровне отдельных видов. Для всех видов, представленных ≥10 живыми деревьями в каждой категории, сравнивались диаметр и объём ствола с использованием теста Манна–Уитни. Структура запаса древесины по видам анализировалась на основе долей участия каждого вида в общем запасе на 1 га; для оценки различий между категориями участков применялся χ²-критерий.
Первичная обработка данных проводилась в среде Excel. Карта типов леса была составлена на основе цифровой базы лесоустройства, пространственный анализ выполнен в QGIS. Все дальнейшие расчёты и визуализация выполнялись в среде Python с использованием следующих библиотек: pandas (McKinney, 2010), numpy (Harris et al., 2020), scipy (Virtanen et al., 2020), matplotlib (Hunter, 2007), seaborn (Waskom, 2021).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Видовой состав. На старовозрастных участках было отмечено 19 видов деревьев. Наиболее многочисленными по числу стволов оказались пихта белокорая (Abies nephrolepis (Trautv.) Maxim.), ель аянская (Picea ajanensis Fisch. ex Carrière), липа амурская (Tilia amurensis Rupr.), кедр корейский (Pinus koraiensis Sieb. et Zucc.), берёза ребристая, или жёлтая (Betula costata Trautv.), клён жёлтый (Acer ukurunduense Trautv. et C.A. Mey.), клён мелколистный (Acer mono Maxim. ex Rupr.). Также присутствовали дуб монгольский (Quercus mongolica Fisch. ex Ledeb.), вяз, или ильм лопастной (Ulmus laciniata (Trautv.) Mayr), тополь Максимовича (Populus maximowiczii A. Henry), бархат амурский (Phellodendron amurense Rupr.), клён зеленокорый (Acer tegmentosum Maxim. et Rupr.) и вишня Максимовича (Cerasus maximowiczii (Rupr.) Kom.). Единично встречались берёза плосколистная, или маньчжурская (Betula platyphylla Sukaczev), ель корейская (Picea koraiensis Nakai), рябина амурская (Sorbus amurensis Koehne), сирень амурская (Syringa amurensis Rupr.), тополь дрожащий, черемуха Маака (Padus maackii (Rupr.) Kom.) и ясень маньчжурский (Fraxinus mandshurica Rupr.).
На послерубочных участках было отмечено 25 древесных видов. Видовой состав в целом перекрывает состав, характерный для старовозрастных участков, однако дополняется рядом пионерных или светолюбивых видов. Помимо массовых видов, общих для обеих категорий, в составе послерубочных древостоев значительное участие принимают берёза маньчжурская, тополь Максимовича и тополь дрожащий (Populus tremula L.), ложнотополь сердцелистный, или ива сердцелистная (Salix cardiophylla Trautv. et C.A. Mey), ольха пушистая (Alnus hirsuta (Spach) Turcz. ex Rupr.), бархат амурский, ива козья (Salix caprea L.), ива росистая (Salix rorida Laksch.) и жестер даурский, или крушина даурская (Rhamnus davurica Pall.). Интересно, что тис остроконечный (Taxus cuspidata Sieb. et Zucc.) был отмечен только на послерубочных участках. Это, с одной стороны, связано с его редкостью, и, с другой стороны, более успешным возобновлением после рубок. Некоторые виды, обычные в старовозрастных древостоях, почти полностью исчезают в послерубочных лесах – особенно нужно отметить кедр корейский. Это указывает на нарушение устойчивой структуры коренного сообщества и его частичном замещении временными сукцессионными элементами.
Общие характеристики. На старовозрастных участках всего зафиксировано 2534 дерева, а на послерубочных – 2037. Густота древостоя (таблица) при пересчёте на 1 га на старовозрастных участках несколько выше, чем на послерубочных. Суммарная площадь поперечного сечения стволов на гектар также выше на старовозрастных участках. Средняя высота деревьев практически не отличается, но стандартное отклонение выше на старовозрастных участках, что свидетельствует о большей вертикальной неоднородности. Средний диаметр деревьев на старовозрастных участках составляет 20.6 см при SD = 15.1 см, на послерубочных – 19.3 см при SD = 13.4 см. Средний объём одного ствола выше на старовозрастных участках (0.54 м³ против 0.43 м³), с большей дисперсией (SD = 0.99 м³ против 0.82 м³). Особенно нужно отметить различия в запасе древесины, которые составляют 116 м³га-1 (или 106 м³га-1, если учитывать только живые деревья). Таким образом, старовозрастный древостой не только характеризуется более высокой продуктивностью, но и большей гетерогенностью структуры древостоя по большинству параметров.
Таблица. Характеристики древостоя старовозрастных и послерубочных участков
| Показатель, единица измерения | Старовозрастные участки | Послерубочные участки |
| Число деревьев на 1 га, шт. | 918 | 878 |
| Средний диаметр (DBH), см (SD) | 20.6 (15.1) | 19.3 (13.4) |
| Средняя высота, м (SD) | 16.1 (5.8) | 16.2 (5.4) |
| Средний объём ствола, м³ (SD) | 0.54 (0.99) | 0.43 (0.82) |
| Медианный диаметр, см (25%, 75% перцентиль) | 15.2 (10.1, 26.5) | 15.0 (9.8, 24.5) |
| Медианная высота, м (25%, 75% перцентиль) | 15.6 (10.5, 20.2) | 15.5 (10.4, 19.8) |
| Медианный объём ствола, м³ (25%, 75% перцентиль) | 0.16 (0.026, 0.55) | 0.14 (0.024, 0.45) |
| BA на 1 га, м² | 47.1 | 38.0 |
| Запас древостоя на 1 га, м³, живые; сухостой | 496; 38 | 380; 28 |
Примечания: BA – сумма площадей поперечных сечений, SD – стандартное отклонение.
Полученные медианы диаметра на высоте груди (DBH), объёма ствола и высоты оказались практически одинаковыми для старовозрастных и послерубочных участков. Такая близость медианных значений указывает на то, что «типичное» дерево в послерубочном древостое по своим размерам почти не отличается от дерева в старовозрастном. Однако в старовозрастных древостоях существенно выше доля крупных деревьев, что отражается в более широких межквартильных интервалах. Таким образом, медианы не выявляют различий, связанных с присутствием крупных экземпляров, но хорошо отражают сходство массовых классов стволов в обеих категориях участков.
Распределение диаметров и объемов стволов. Результаты анализа распределения диаметра и объёма ствола представлены в гистограммах (рис. 2, 3). В старовозрастном древостое распределение диаметра деревьев более широкое и равномерное: представлены все ступени толщины, включая крупномерные деревья с диаметром более 40 см. В послерубочном древостое преобладают деревья с диаметром до 20 см; более крупные экземпляры почти не встречаются. Аналогичная картина наблюдается в распределении по объёму ствола: в старовозрастном древостое встречаются деревья с объёмом до 2–3 м³ и выше, в то время как в послерубочном доминируют экземпляры с объёмом менее 0.5 м³. Вклад деревьев со значительным запасом в послерубочном древостое минимален.
Различия в форме распределений подчёркивают ключевое отличие между двумя категориями древостоев. Старовозрастные участки характеризуются более сложной структурой с выраженной размерной и возрастной дифференциацией, тогда как послерубочные остаются относительно однородными по составу и структуре.
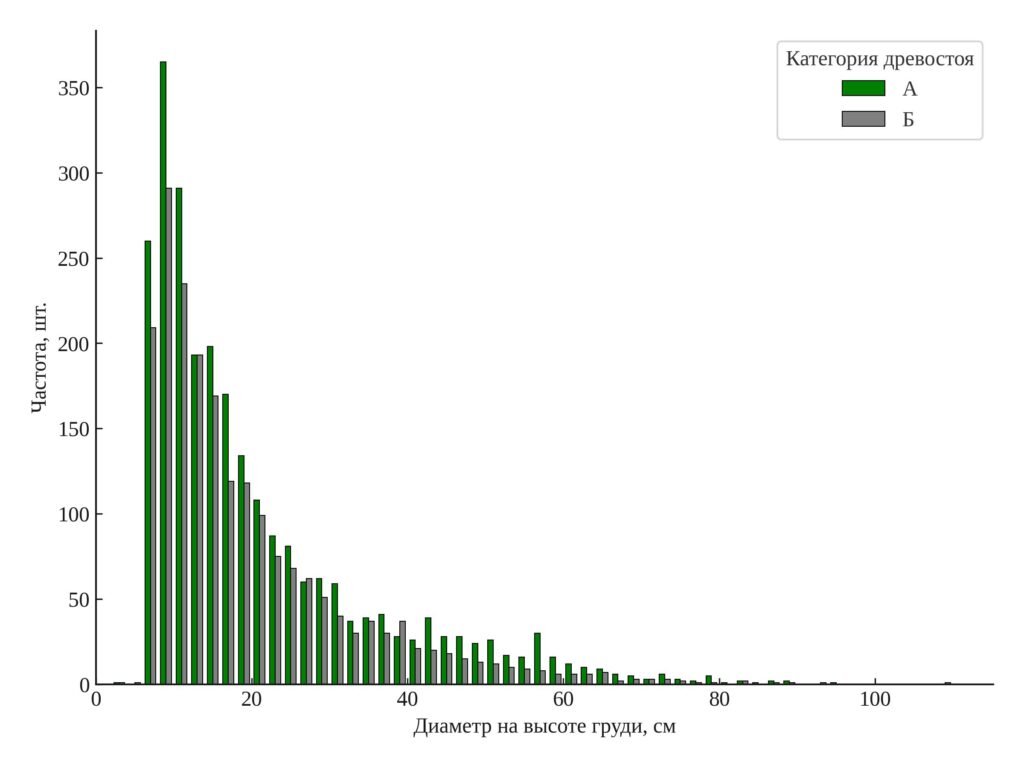
Рисунок 2. Распределение деревьев по диаметру (DBH). А — старовозрастный древостой, Б — послерубочный древостой
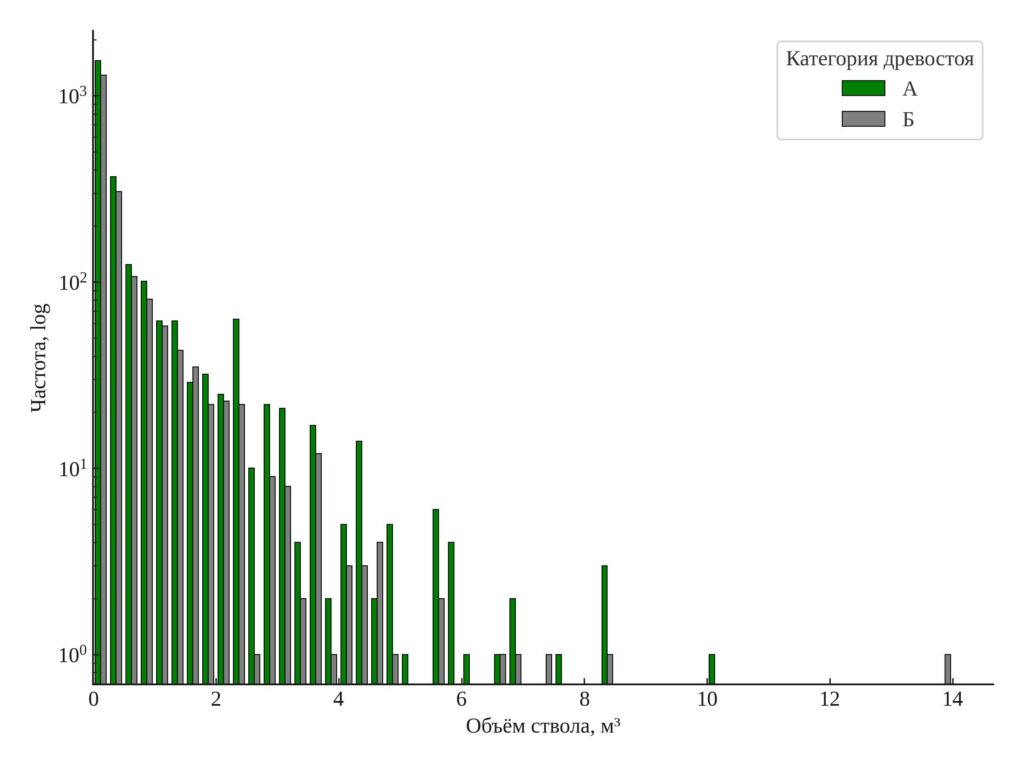
Рисунок 3. Распределение деревьев по запасу (ось ординат — логарифмическая). А — старовозрастный древостой, Б — послерубочный древостой
Сравнение распределений основных признаков деревьев. Результаты непараметрического теста Колмогорова–Смирнова (рис. 4) демонстрируют статистически значимые различия в распределениях всех исследованных признаков между старовозрастными и послерубочными участками. Наибольшее отличие выявлено по высоте (KS = 0.057, p = 0.0012), что указывает на неоднородность вертикальной структуры. Существенные различия также установлены для диаметра (KS = 0.043, p = 0.0285), суммы площадей сечения стволов (KS = 0.043, p = 0.0285) и объёма ствола (KS = 0.047, p = 0.0124). Это подтверждает, что даже спустя 60–80 лет после нарушений послерубочный древостой сохраняет упрощённую структуру и не достигает пространственной и размерной сложности, характерной для старовозрастного древостоя.
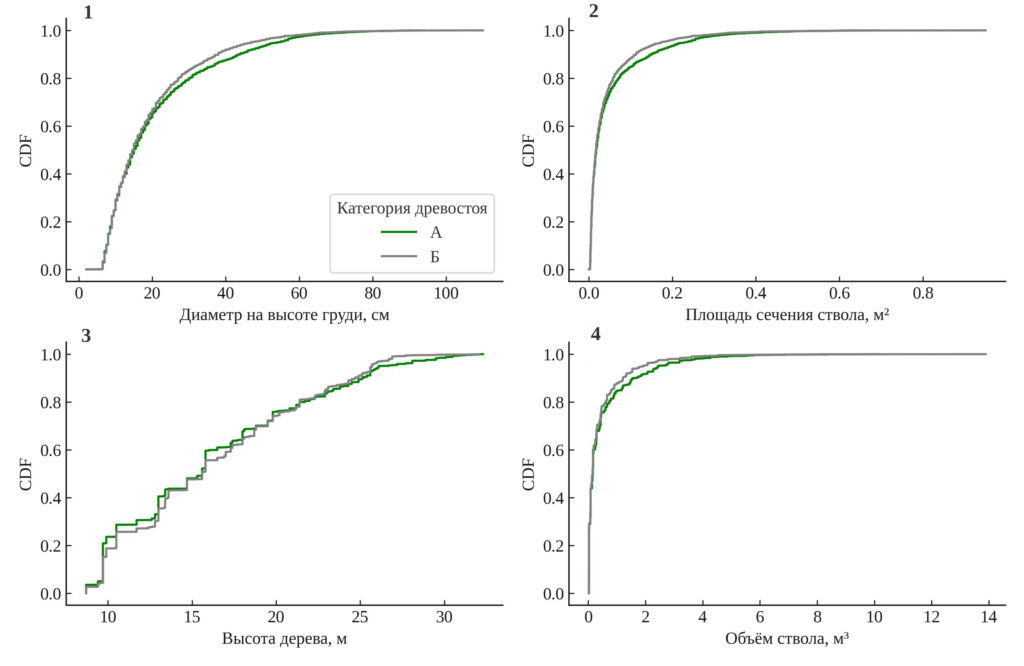
Рисунок 4. Функции распределения (CDF) основных признаков деревьев. 1 — диаметр на высоте груди (DBH), см; 2 — сумма площадей поперечных сечений, м²; 3 — высота дерева, м; 4 — объём ствола, м³. А — старовозрастный древостой, Б — послерубочный древостой
Характеристика отдельных видов. Рисунок 5 иллюстрирует различия в диаметре на высоте груди и объёме ствола у древесных видов, произрастающих на старовозрастных и послерубочных участках. Из анализа были исключены ель корейская и тис остроконечный, поскольку деревьев этого вида слишком мало для расчёта средних величин и их вариации. Средние значения диаметра ствола и объёма древесины на один ствол существенно варьируют между древесными видами, а также между старовозрастными и послерубочными участками. Наибольшие размеры стволов характерны для кедра корейского и ели аянской: у кедра средний диаметр на старовозрастных участках составляет 40.9 см, а на послерубочных – 24.5 см, что на 40% меньше. Аналогичная тенденция наблюдается и для объёма ствола: 2.42 м³ против 1.09 м³ соответственно, разница составляет 55%. У ели аянской средний диаметр уменьшается с 26.2 до 21.7 см (−17%), а объём – с 0.69 до 0.46 м³ (−33%).
Сходные значения диаметров наблюдаются у липы амурской и дуба монгольского в обеих категориях древостоев. Так, у липы средний диаметр составляет 23.2 см в старовозрастных лесах и 22.5 см в послерубочных, а объём — 0.55 и 0.47 м³ соответственно. У берёзы жёлтой, напротив, наблюдается резкое снижение диаметра с 42.0 до 22.0 см (−48%), а объёма — с 1.66 до 0.48 м³ (−71%).
Во многих случаях стандартные отклонения также выше в старовозрастных лесах, что свидетельствует о большей неоднородности и возрастной мозаичности древостоя. Для некоторых видов данные доступны только по одной из категорий, что связано с их отсутствием или единичными представителями в другой. Это касается, в частности, бархата амурского, ильма долинного и ивы козьей, которые встречаются исключительно на послерубочных участках.
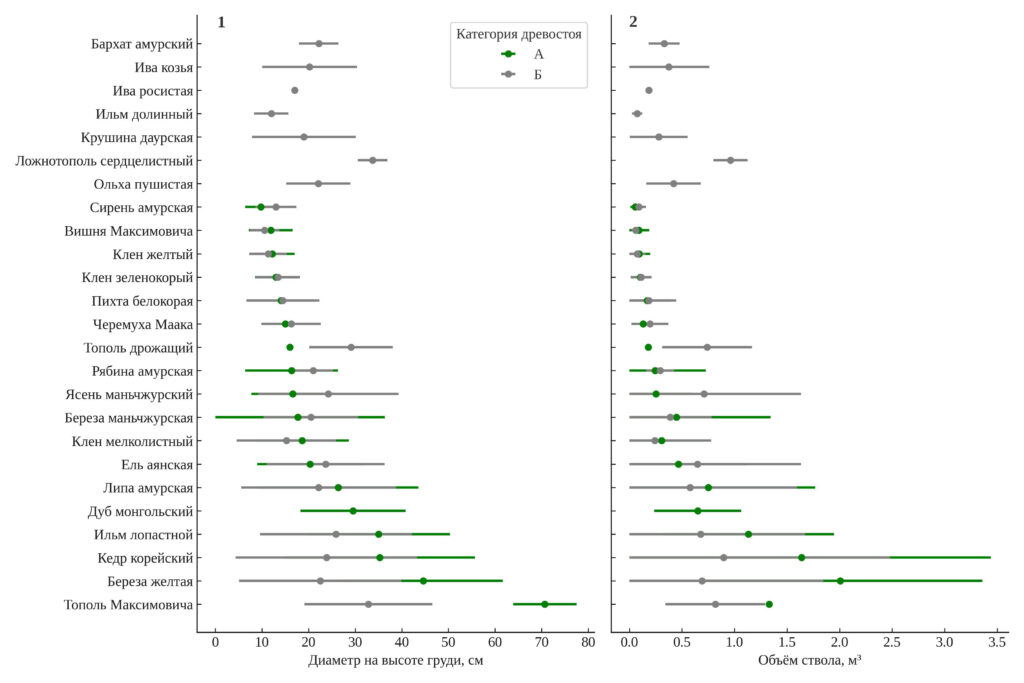
Рисунок 5. Средние значения диаметра (DBH) (1) и объёма ствола (2) деревьев различных видов в старовозрастных (А) и послерубочных древостоях (Б) (± SD)
Для оценки различий в морфометрических признаках деревьев (диаметр на высоте груди и объём ствола) между старовозрастными и послерубочными участками применялся непараметрический тест Манна–Уитни. В анализ включались только те виды, для которых в каждой категории насчитывалось не менее 10 живых деревьев (всего 11 видов).
Сравнение медианных значений диаметра на высоте груди показало статистически значимые различия для всех пяти наиболее массовых видов: берёзы жёлтой, кедра корейского, липы амурской, клёна мелколистного и ели аянской (p < 0.01 во всех случаях). У первых четырёх видов диаметр деревьев был существенно выше на старовозрастных участках, в то время как для ели аянской наблюдалась обратная тенденция — более крупные деревья встречались на послерубочных участках. Аналогичная закономерность выявлена и по объёму ствола: у берёзы жёлтой и кедра медианный объём на старовозрастных участках в несколько раз превышал значения на послерубочных. Таким образом, структура древостоя по ряду ключевых видов существенно различается между категориями.
Характеристика накопления запаса в старовозрастных и нарушенных лесах. Рисунок 6 иллюстрирует распределение запаса живой древесины между видами на старовозрастных и послерубочных участках. На старовозрастных участках доминирует кедр корейский, обеспечивая 32.0% общего запаса (146.6 м³га-1). Его вклад в запас послерубочных участков снижается более чем в девять раз — до 4.7% (16.6 м³га-1), что указывает на слабую восстановительную динамику этого вида после нарушений. Ель аянская и берёза жёлтая, напротив, показывают высокие значения запаса в категориях: их суммарный вклад составляет 29.1% на старовозрастных и 42.7% на послерубочных участках, что отражает их устойчивость к нарушениям и способность формировать как стабильные, так и вторичные сообщества.
В послерубочных древостоях заметно увеличивается доля пионерных видов: тополя Максимовича (5.3% запаса), тополя дрожащего (2.6%), черёмухи Маака (0.8%), ольхи пушистой (0.8%) и берёзы маньчжурской (2.0%). В старовозрастных древостоях они почти не участвуют в формировании запаса. Эти виды, наряду с другими мелколиственными и светолюбивыми видами, формируют характерную структуру ранней стадии сукцессии. Таким образом, в результате рубок происходит смещение в сторону быстрорастущих видов, а участие кедра и других позднесукцессионных видов существенно снижается.
Для оценки различий в структуре запасов древесины между старовозрастными и послерубочными участками был проведён χ²-тест, основанный на долях участия древесных видов в суммарном запасе на 1 га. Для каждого вида был рассчитан его вклад в общий запас в пределах каждой категории участков.
Результаты теста показали, что распределение запасов по видам между двумя категориями достоверно различается (χ² = 150.66; p < 0.0001). Это подтверждает вывод о том, что вклад отдельных видов в общий запас изменяется как по величине, так и по составу. В целом, структура доминирования в послерубочных древостоях формируется за счёт более быстрых и пластичных видов, в то время как участие коренных видов существенно снижается.
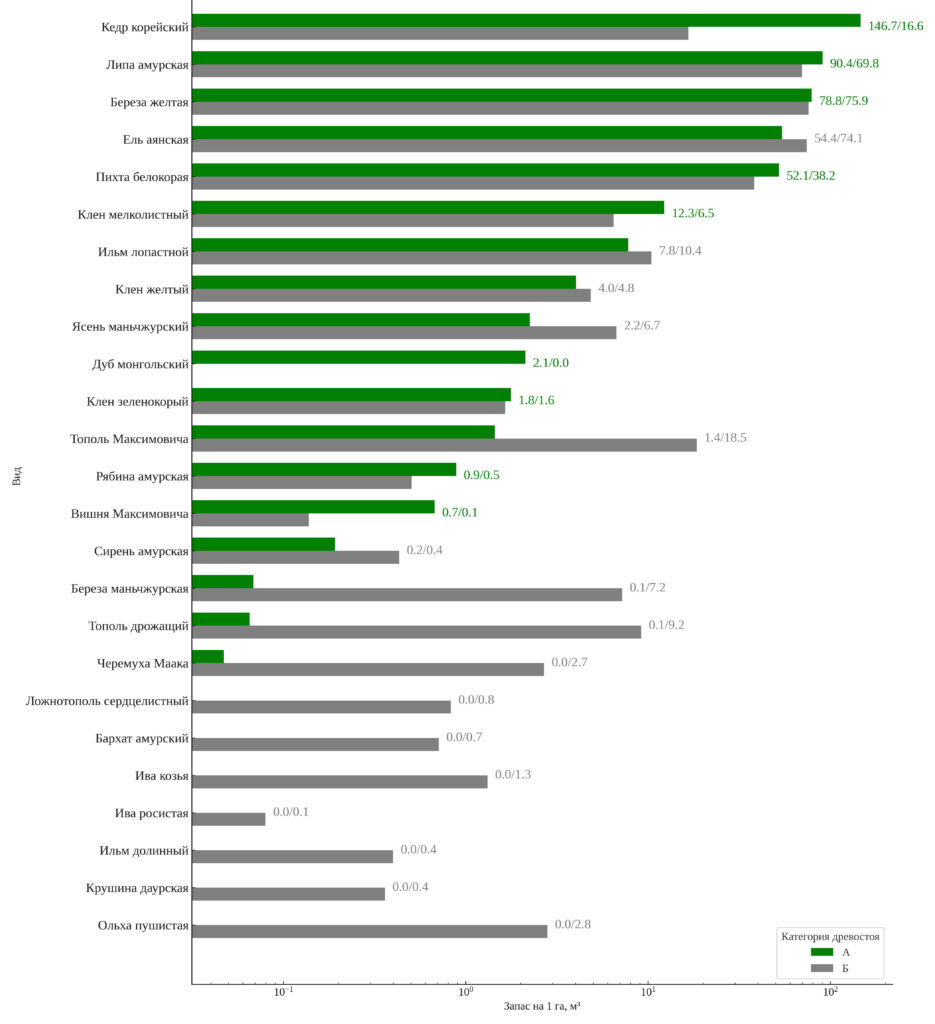
Рисунок 6. Запас древесины на 1 га по видам в старовозрастных (А) и послерубочных (Б) древостоях
Состояние древостоя через 60-70 лет после рубки и возможность восстановления его исходного состояния. Восстановление кедрово-широколиственных лесов после промышленных рубок представляет собой сложный и медленный процесс, зависящий от многих биотических и абиотических факторов, что подчёркивается в ряде исследований, включая обзор Манько и др. (2009) и аналитические данные Ковалева и Качановой (2023), указывающих на длительность и неоднозначность восстановления даже после однократных рубок. Оценка состояния послерубочных древостоев через 60–70 лет после однократных рубок позволяет судить о направлении и степени сукцессионных изменений. В нашем исследовании, основанном на данных, собранных в сопоставимых условиях в южном Сихотэ-Алине, выполнено сравнение древостоев, подвергшихся рубкам в 1960-х годах, со старовозрастными ненарушенными древостоями. Это даёт возможность оценить степень сближения структуры лесов после рубок с исходной и выявить признаки, свидетельствующие о сохраняющейся трансформации. Нужно отметить, что в рассматриваемом случае рубки носили приисковый характер: велась заготовка исключительно кедра корейского и ели аянской, при этом изымались единичные крупномерные деревья, а вмешательство в структуру древостоев было минимальным.
Оценка таких показателей, как средний диаметр, высота, сумма площадей сечения и запас выявила характерные различия между послерубочными и старовозрастными древостоями. На послерубочных участках преобладают деревья с меньшим диаметром: средний диаметр (19.3 см против 20.6 см) и, особенно, запас на 1 га (380 м³га-1 против 496 м³га-1) ниже по сравнению с ненарушенными участками. Гистограммы распределения диаметра и объёма стволов демонстрируют отсутствие крупных деревьев в послерубочных древостоях, тогда как в старовозрастных древостоях представлены все классы, включая единичные экземпляры с объёмом ствола до 10–14 м³. Эти различия статистически значимы, что подтверждается результатами теста Колмогорова–Смирнова.
Подобная ситуация описана в исследованиях темнохвойных лесов после сплошных рубок. Например, по данным Лихановой и др. (2021), в черничных ельниках средней тайги через 50 лет после рубки восстанавливается не более 50–60% исходного состава и структуры. При этом авторами подчёркивается, что даже спустя 60 лет структура фитоценозов остаётся существенно отличной от коренной, а восстановление полноразмерного древостоя требует значительно более длительного времени.
Полученная в данном исследовании картина полностью согласуется с этими выводами: временной промежуток в 60–70 лет после однократной рубки недостаточен для восстановления исходной размерной структуры кедрово-широколиственных лесов.
Существенные различия между послерубочными и старовозрастными участками выявлены и в видовом составе древостоев. На участках, не подвергавшихся рубке, основным лесообразующим видом остаётся кедр корейский, вносящий наибольший вклад в общий запас древостоя. В то же время на послерубочных участках доля кедра резко снижена, а доминирующее положение занимают быстрорастущие лиственные виды – берёза, тополя, черёмуха и ольха.
Сходные закономерности описаны Ковалевым и соавт. (2021), где подчёркивается, что после рубок кедр долго не занимает лидирующее положение в древостое, уступая его быстрорастущим лиственным видам, и даже при наличии подроста темпы восстановления кедра недостаточны для его участия в формировании древостоя. В более поздней работе Ковалёва и Качановой (2023) указывается, что полноценное восстановление структуры кедровников после рубки может занять более 150 лет, и без активных мер по содействию возобновлению кедра его участие в древостоях остаётся крайне ограниченным. Авторы подчёркивают, что в текущих условиях хозяйственного использования такие сроки почти недостижимы. Аналогичные выводы сделаны в исследованиях А. И. Кудинова (2012, 2014), основанных на 40-летнем мониторинге послерубочных участков в южном Приморье: несмотря на сохранение отдельных хвойных элементов в подросте, в древостоях доминируют лиственные виды. Автор предполагает, что даже при наличии подроста кедра и потенциальной возможности его участия в формировании древостоя, структура сообщества остаётся стабильной и ориентированной на преобладание лиственных видов. Смена господства лиственных пород на кедр корейский совершается в течение 120-160 лет после рубки и только при наличии предварительного подроста в количестве 400–500 экз. га-1. В работе C. Г. Глушко с соавторами (2022), посвящённой сукцессионной динамике пихтово-еловых лесов, подчёркивается, что восстановление исходной структуры происходит неравномерно и с высокой пространственной мозаичностью, особенно в отсутствие целенаправленных лесохозяйственных мероприятий.
Региональные обзоры (Манько и др., 2009; Манько, Петропавловский, 2010) подтверждают, что трансформация кедровников в производные леса после хозяйственного вмешательства имеет повсеместный характер, а восстановление их исходной структуры может занимать столетия. Формирующиеся сообщества характеризуются устойчивой динамикой, часто с уклоном в сторону альтернативных типов леса – дубрав, ельников, кленово-липовых и смешанных лиственных насаждений. Эти изменения сопровождаются снижением продуктивности, биологического разнообразия и защитных функций.
Сходные результаты получены и для темнохвойных лесов Европейской России. По данным И. А. Лихановой с соавторами (2021), даже через 50–60 лет после сплошных рубок восстановление исходной структуры происходит не полностью: сукцессия нередко завершается формированием стабильных вторичных сообществ с иным доминирующим составом. Аналогичная ситуация наблюдается и в умеренных лесах Скандинавии: Asplund et al. (2020) показали, что вторичные леса сохраняют отличия в видовом составе, вертикальной структуре и составе подлеска даже спустя полвека после рубок. Авторы подчёркивают устойчивость так называемого «наследия лесопользования», которое формирует долгосрочную, но отличную от исходной структуру лесного сообщества, требующую либо целенаправленного вмешательства, либо длительного времени для восстановления. Для кедрово-широколиственных лесов юга российского Дальнего Востока это явление не столь выражено, однако сроки восстановления значительно превышают оборот рубок (Ковалёв, Качанова, 2023).
Обобщение литературных данных (Кудинов, 2012; Кудинов, 2014; Ковалев и др., 2021, 2023; Манько и др., 2009) и полученных в данном исследовании результатов позволяет заключить: даже при однократном и давно прошедшем вмешательстве восстановление исходной структуры кедрово-широколиственных лесов протекает крайне медленно и остаётся незавершённым даже спустя 60–70 лет. Послерубочные древостои существенно отличаются от старовозрастных по запасу древесины, размерной структуре и доминирующим видам. В них отсутствуют крупные деревья, преобладают светолюбивые лиственные породы, а участие кедра минимально.
Такие сообщества представляют собой переходную сукцессионную стадию, на которой пионерные виды ещё сохраняют доминирующее положение, тогда как позднесукцессионные – кедр корейский, ель аянская, берёза жёлтая – начинают накапливать запас. Это свидетельствует о высокой инерционности нарушенных кедрово-широколиственных лесов и длительных последствиях даже слабых воздействий, подчёркивая необходимость пересмотра подходов к их охране, восстановлению и лесопользованию.
ЗАКЛЮЧЕНИЕ
Проведённое исследование показало, что спустя 60–70 лет после однократных выборочных рубок приискового характера структура и состав древостоев остаются существенно отличными от старовозрастных кедрово-широколиственных лесов. Послерубочные древостои характеризуются меньшим запасом древесины, значительным участием пионерных лиственных видов и отсутствием крупных деревьев, что свидетельствует о незавершённости процессов восстановления. Хотя кедр корейский сохраняется в составе, он по-прежнему не восстанавливает доминирующего положения: его участие в структуре и темпы роста остаются недостаточными для конкуренции с быстрорастущими лиственными видами.
Таким образом, восстановление исходной структуры кедрово-широколиственных лесов представляет собой медленный и нестабильный процесс, зависящий от множества факторов и требующий продолжительного времени. Даже при минимальном вмешательстве возврат к коренному состоянию может занимать более 100 лет и, как правило, не происходит в пределах одного оборота рубок без активного содействия со стороны лесоводства.
БЛАГОДАРНОСТИ
Работа выполнена в рамках государственного задания Министерства науки и высшего образования Российской Федерации (тема № 124012400285-7), а также в рамках проекта по разработке системы наземного и дистанционного мониторинга пулов углерода и потоков парниковых газов на территории Российской Федерации, создания системы учета данных о потоках климатически активных веществ и бюджете углерода в лесах и других наземных экологических экосистемах (1 этап).
СПИСОК ЛИТЕРАТУРЫ
Архив ДВО РАН, отчёты ВУС 1964–1965 гг.
Громцев А. Н. Ландшафтная экология таежных лесов: теоретические и прикладные аспекты. Петрозаводск: Карельский научный центр РАН, 1999. 144 с.
Глушко С. Г., Комарова Т. А., Прохоренко Н. Б. Формирование молодняков после рубок и пожаров в темнохвойных осоково-папоротниковых лесах Южного Сихотэ-Алиня // Лесоведение. 2022. № 2. С. 144–156.
Гуков Г. В. Лесоводство Дальнего Востока. Владивосток: Изд-во Дальневост. ун-та, 1989. 215 с.
Данилин А. К., Чумин В. Т. Леса и лесное хозяйство Хабаровского края. Хабаровск: Кн. Изд-во, 2000. 416 с.
Лиханова И. А., Перминова Е. М., Шушпанникова Г. С., Железнова Г. В., Пыстина Т. Н., Холопов Ю. В. Динамика растительности после сплошнолесосечных рубок ельников черничных (среднетаежная подзона Европейского северо-востока России) // Растительность России. 2021. № 40. С. 108–136. DOI: 10.31111/vegrus/2021.40.108
Ковалев А. П. Эколого-лесоводственные основы рубок в лесах Дальнего Востока. Хабаровск: ДальНИИЛХ, 2004. 267 с.
Ковалев А. П., Качанова Т. Г. О будущем кедрово-широколиственных лесов Дальнего Востока // Лесное хозяйство и лесная промышленность. 2023. № 1. С. 10–12.
Кожевникова Н. К. Динамика погодно-климатических характеристик и экологические функции малого лесного бассейна // Сибирский экологический журнал. 2009. № 5. С. 693–703.
Колесников Б. П. Кедровые леса Дальнего Востока // Труды Дальневосточного филиала им. В.Л. Комарова. Серия ботаники АН СССР. 1956. Т. 2 (4). 262 с.
Комарова Т. А., Прохоренко Н. Б., Глушко С. Г., Терехина Н. В. Лесовосстановительные сукцессии после рубок главного пользования в кедрово-пихтово-еловых широкотравно-осоково-папоротниковых лесах Южного Сихотэ-Алиня // Ботанический журнал. 2022. Т. 107, № 10. С. 939–953.
Кудинов А. И. Широколиственно-кедровые леса Южного Приморья и их динамика: автореф. дис. … доктора биол. наук. Владивосток, 2007. 24 с.
Кудинов А. И. Динамика производных фитоценозов на юге Приморского края. Владивосток: Дальнаука, 2012. 140 с.
Кудинов А. И. Послерубочная динамика хвойно-широколиственных лесов южного Приморья // Лесоведение. 2014. № 3. С. 39-48.
Куренцова Г. Э. Динамика кедровых лесов Приморья. Владивосток: ДВО АН СССР, 1968. 132 с.
Лебедев А. А. Лесоводственные мероприятия в кедровниках. СПб.: Лань, 2003. 208 с.
Манько Ю. И., Кудинов А. И., Гладкова Г. А., Бутовец Г. Н. Грабовый широколиственно-елово-кедровый лес за период 1926–2003 гг. (Уссурийский заповедник, Южное Приморье) // Сибирский экологический журнал. 2009. № 6. С. 917–926.
Манько Ю. И., Петропавловский Б. С. Пути оптимизации охраны и использования лесов Приморского края в современных условиях // Проблемы лесоведения и лесоводства: Материалы Всерос. конф. «Мелеховские чтения». Архангельск, 2010. С. 175–178.
Розенберг В. А. Классификация рубок в кедровниках // Лесное хозяйство. 1975. № 4. С. 34–38.
Сибирина Л. А., Гладкова Г. А., Омелько А. М., Баркалов В. Ю. Верхнеуссурийский биогеоценотический стационар Федерального научного центра биоразнообразия наземной биоты Восточной Азии ДВО РАН // Вестник ДВО РАН. 2022. № 4. С. 84–100. DOI: 10.37102/0869-7698_2022_224_04_8.
Соловьев К. П. Некоторые факторы, влияющие на возобновление кедра под пологом леса и на лесосеках после выборочных и сплошных рубок // Сборник трудов ДальНИИЛХ. 1948. Вып. 1. С. 58–107.
Усенко Н. В. Деревья, кустарники и лианы Дальнего Востока. 2-е изд., перераб. и доп. Хабаровск, 1984. 272 с.
Яковлева А. Н. Эколого-фитоценотические особенности пространственного распределения лесной растительности Южного Сихотэ-Алиня: на примере Верхнеуссурийского стационара: Дисс. … канд. биол. наук (спец. 03.00.05, 03.00.16). Владивосток: БПИ, 2004.
Asplund J., Nordén J., Kjønaas O. J., Madsen R. L., Lunde L. F., Birkemoe T., … & Nybakken L. Long term effects of forest management on forest structure and dead wood in mature boreal forests // Forest Ecology and Management. 2024. Vol. 572. Article 122315. DOI: 10.1016/j.foreco.2024.122315.
Harris C. R., Millman K. J., van der Walt S. J., Gommers R., Virtanen P., Cournapeau D., … & Oliphant T. E. Array programming with NumPy // Nature. 2020. Vol. 585. P. 357–362.
Hunter J. D. Matplotlib: A 2D graphics environment // Computing in Science & Engineering. 2007. Vol. 9, No. 3. P. 90–95.
McKinney W. Data Structures for Statistical Computing in Python // Proceedings of the 9th Python in Science Conference. 2010. P. 51–56.
Virtanen P., Gommers R., Oliphant T. E., Haberland M., Reddy T., Cournapeau D., … & Van Mulbregt P. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python // Nature Methods. 2020. Vol. 17. P. 261–272.
Waskom M. L. Seaborn: statistical data visualization // Journal of Open Source Software. 2021. Vol. 6, No. 60. Article 3021. DOI: 10.21105/joss.03021
Рецензент: к. б. н., в. н. с. Коротков В. Н.





