- DOI: 10.31509/2658-607x-202471-141
- УДК 574.472
Роль эктомикоризы древесных растений в динамике органического вещества лесных почв: обобщение и концептуализация имеющихся данных
![]()
© 2024 г. О. Г. Чертов1*, И. В. Припутина2, В. Н. Шанин1, 2, П. В. Фролов2
1Центр по проблемам экологии и продуктивности лесов РАН,
Россия, 117997, г. Москва, ул. Профсоюзная, 84/32, стр. 14
2Институт физико-химических и биологических проблем почвоведения
РАН — обособленное подразделение Федерального исследовательского центра
«Пущинский научный центр биологических исследований Российской академии наук»
Россия, 142290, Московская обл., г. Пущино, ул. Институтская, д. 2, корп. 2.
*E-mail: ochertov@rambler.ru
Поступила в редакцию: 20.02.2024
После рецензирования: 15.03.2024
Принята к печати: 24.03.2024
Рассмотрены основные аспекты функционирования эктомикоризы древесных растений, которые влияют на формирование и разложение почвенного органического вещества (ПОВ) лесных почв. Эктомикориза потребляет на свой рост корневые экссудаты деревьев и образует густую сеть внешнего мицелия, который имеет короткий жизненный цикл. Благодаря этому происходит быстрый оборот биомассы мицелия с формированием большого количества внутрипочвенного опада, превышающего по массе поверхностный опад листвы/хвои. Био- и некромасса мицелия потребляется почвенной биотой разных функциональных групп и уровней почвенных пищевых сетей (soil food webs) с образованием твердофазных продуктов метаболизма, представляющих собой лабильный богатый азотом пул молодого органического вещества почвы. Это определяет весомый вклад эктомикоризы в пополнение ПОВ. Одновременно разложение этого лабильного пула ПОВ служит механизмом возврата в почву азота, доступного для корней растений, что определяет положительный эффект эктомикоризы на рост деревьев и других растений в лесных сообществах. Таким образом, рассмотрение процессов влияния эктомикоризы на органическое вещество почвы должно включать триаду организмов “растение – микориза – почвенная биота” (включая как микроорганизмы, так и почвенную фауну).
Ключевые слова: лесные почвы, эктомикориза, органическое вещество почв, майнинг азота, почвенная фауна, экскреты фауны
Взаимодействие растения и почвы в экологии обычно рассматривается в рамках концепции биологического круговорота: почва снабжает растения элементами питания и влагой, растение возвращает в почву с опадом вещество и энергию, включая органическое вещество и элементы питания (Одум, 1986; Базилевич, 2008; Титлянова, Самбуу, 2016). Однако в природе имеются более тонкие механизмы взаимодействия в системе «растение-почва», в частности: (а) активизация растениями почвенных микроорганизмов в ризосфере (прайминг-эффект) (Blagodatskaya, Kuzyakov, 2008; Bastida, 2019); (б) симбиоз корней растений с микоризными грибами (Воронина, 2006; Smith, Read, 2008; Смит, Рид, 2012); (в) симбиоз корней с азотфиксирующими микроорганизмами (Волобуева, 2011; Mus et al., 2016; Aasfar, 2021; Threatt, Rees, 2023); (г) поступление в почву элементов питания из древесных растений с кроновыми водами (Аккумуляция углерода…, 2018). При этом микоризные грибы являются облигатными биотрофами, существующими за счёт вещества и энергии растения-хозяина (Hobbie et al., 2013). Считается, что совокупный эффект перечисленных выше механизмов приводит к оптимизации среды обитания растений.
Микоризный симбиоз, то есть взаимодействие почвенных грибов с корнями растений, привлекает пристальное внимание исследователей со второй половины XIX в. до настоящего времени. Так, например, обстоятельная монография С. Смит и Д. Рид (2012), посвящённая вопросам изучения микоризы, насчитывает 787 страниц текста, а библиография занимает 120 страниц петитом. Тем не менее, поток публикаций по микоризному симбиозу не снижается (Martin, 2017; Tedersoo, 2017; Brundrett, Tedersoo, 2018; Averill et al., 2019; Becquer et al., 2019; Tedersoo et al., 2020; Hawkins et al., 2023; Bönisch. 2024; и др.). В отечественной литературе заметен теоретический интерес к микоризной тематике (Воронина, 2007; Тиунов, 2014; Макаров, 2019), к описанию микориз различных растений и сообществ (Весёлкин, 2015; Весёлкин и др., 2015; Колмаков, 2016; Лавренов, 2017; Дудка, 2021; Betekhtina, Veselkin, 2019) и к роли микориз в лесном и сельском хозяйстве (Иозус, 2019; Лукина и др., 2019) с акцентом на микоризу культурных растений (Вацке, 2005; а также большинство из 3800 русскоязычных публикаций в базе данных по микоризе в eLibrary). Более того, грибы в целом и микориза в частности всё чаще попадают в поле зрения научной общественности (Хофрихтер, 2021; Сергеева, 2022), что связано не только с научно-познавательной, но и с практической значимостью этого явления.
Следует подчеркнуть, что микоризный симбиоз широко распространен в растительном покрове планеты (Soudzilovskaia et al., 2017), характеризуется необычайным биоразнообразием грибов (Смит, Рид, 2012; Johnson et al., 2015) и представляет собою достаточно весомый и активный пул углерода в биосфере (Akhmetzhanova et al., 2012; Hawkins et al., 2023). В настоящее время различают следующие типы микориз: эктомикориза в основном древесных растений бореального и умеренного пояса; арбускулярная микориза также древесных и травянистых растений; эрикоидная микориза кустарников семейства Вересковые; орхидная микориза эпифитной растительности тропических лесов, а также смешанная микогетеротрофная (Смит, Рид, 2012). Несомненно, количество этих групп будет возрастать по мере накопления новых данных.
Доминирующими типами в микоризном симбиозе являются экто- и арбускулярная микоризы. Эктомикориза (ЭМ) представляет собою грибной чехол на поверхности тонких активных корневых окончаний растений. Грибница чехла только незначительно проникает в клетки корня (образуя сеть гиф, известную как сеть Гартига), а из чехла растут гифы, формирующие густую сеть внешнего мицелия, который распространяется в почве далеко за пределы ризосферы. Арбускулярная микориза располагается внутри корня, откуда прорастают многочисленные гифы внешнего мицелия, заполняя объём почвы, значительно превышающий тот, который использует собственно корневая система.
ЭМ в лесах отличаются разнообразием (Tedersoo, 2017; Pang et al., 2023) и высокой пространственной вариабельностью по горизонтали и вертикали в масштабе сантиметров в зависимости от видового состава растительности и мозаичности почвенного покрова (O’Hanlon, 2012). Динамика биомассы чехла ЭМ жестко привязана к ростовым окончаниям тонких корней: по мере их роста чехол синхронно “передвигается“ вслед за корневым окончанием, отмирая на покрывающихся корой одревесневших корнях. Биомасса микоризного чехла составляет около 60% учитываемой при полевых исследованиях биомассы тонких корней (Смит, Рид, 2012). Видовой состав ЭМ грибов закономерно меняется в ходе восстановительных сукцессий лесной растительности после различных нарушений (Guignabert et al., 2018; Wang et al., 2023). Привлекает внимание анализ факторов, регулирующих сдвиг равновесия в отношениях между ЭМ грибом и растением-хозяином (Brundrett, Tedersoo, 2018).
Концептуально взаимодействие корней растений и микоризы понятно: микоризный гриб потребляет продукты фотосинтеза растения (углерод органических соединений корневых экссудатов) и расходует их на свой рост. При этом микориза ещё и добывает из органического вещества почвы и её минеральной матрицы те элементы, которых ей не хватает (в хвойных и широколиственных лесах Северного Полушария это в первую очередь азот, а южнее — фосфор). До настоящего времени в литературе доминирует представление о равноправном “обмене” веществом между растением и грибом: растение снабжает микоризу (микоризный гриб) продуктами фотосинтеза для роста мицелия, а микоризный гриб, соответственно, улучшает обеспеченность корней растения-хозяина элементами питания (Fogel, Hunt, 1983; Kranabetter, 2014). Параллельно встречаются работы, рассматривающие взаимодействие растений и микоризы с позиций фитоцентричного (Весёлкин, 2013) или микоцентричного (Staddon, 2005) подходов.
Реальная картина, по-видимому, гораздо сложнее, и до сих пор нет общепринятого объяснения всех механизмов взаимодействия растения и микоризы (Jacquemyn, Merckx, 2019; Stuart, Plett, 2020; Albornoz et al., 2021; Savolainen, Kytöviita, 2022). Значительная часть работ посвящена изучению микоризного континуума мутуализм-паразитизм (Johnson, Graham, 2013), а также анализу факторов, регулирующих сдвиг равновесия в отношениях между грибом и растением (Brundrett, 2004).
В данной работе роль микоризного симбиоза в питании растений и динамике органического вещества лесных почв обсуждается на примере системы “растение – микориза – почвенные микроорганизмы и фауна почвенных пищевых сетей – ПОВ” с акцентом на ЭМ лесов таёжной зоны, которые наиболее распространены в России. Цель работы – краткий анализ актуальных на сегодняшний день представлений о роли ЭМ в лесных экосистемах, а также попытка компиляции определённой непротиворечивой картины функционирования ЭМ с экологической (точнее, экосистемной) точки зрения. Основное внимание уделено процессам, связанным с функционированием внешнего мицелия ЭМ.
Питание микоризы. Корневые экссудаты. Основным источником питания микоризных грибов служат корневые экссудаты (Jones et al., 2004; Смит, Рид, 2012; Pausch et al., 2013; Martin, 2017), представляющие собою продукты фотосинтеза растения-хозяина. В их составе обнаруживается низкомолекулярное органическое вещество: карбоновые кислоты, аминокислоты, моносахариды и комплекс органических веществ разного состава, определяемого общим термином “растворенное органическое вещество” (РОВ) (Kuzyakov et al., 2007; Nehls et al., 2010; Canarini et al., 2019). На долю органических соединений, потребляемой микоризой при разложении ПОВ, приходится незначительная часть: в биомассе микоризного гриба обнаруживается менее 2% углерода, полученного из ПОВ (Talbot et al., 2013; Rineau et al., 2013; Tunlid et al., 2017).
Подчёркивается, что микоризный гриб не является сапротрофным организмом (Nehls et al., 2010; Lindahl, Tunlid, 2015; Nehls et al., 2017; Saifuddin et al., 2021). Действительно, ЭМ не захватывает свежие растительные остатки и древесный отпад, а избирательно разлагает только богатое азотом устойчивое ПОВ (гуминовые кислоты и гумины) (Hobbie et al., 2013). Можно заключить, что микоризные грибы – это облигатные автохтонные организмы, в отличие от остальных сапротрофных грибов и бактерий, которые бывают и зимогенными (разлагающие опад и лабильный ПОВ), и автохтонными.
Микоризные грибы не являются простыми потребителями корневых экссудатов растения-хозяина. Из почвы в корни растения-хозяина за счет микоризного симбиоза одновременно с транспирационным током воды поступают определенные стимуляторы фотосинтеза. В итоге это приводит к заметному увеличению нетто биологической продуктивности (НБП) растения, в результате чего происходит заметное возрастание количества выделяемых корневых экссудатов (Van der Heijden, Horton, 2009; Agerer et al., 2012).
Рост эктомикоризы и продукция внешнего мицелия. По имеющимся оценкам, доля углерода НБП растения-хозяина, забираемая ЭМ, достигает значительных величин, что определяет рост и активность ЭМ (Wallander et al., 2001; Johansson et al., 2008). В целом деревья могут расходовать на функционирование всей корневой системы (корни плюс микориза) до 50% своей НБП (Nehls et al., 2010). На рост собственно микоризы может приходиться до 10-25% от величины НБП растения-хозяина (Nehls et al., 2017), хотя есть и более скромные оценки – до 22% НБП не всего растения, а только части, расходуемой на подземную биомассу (Hobbie, 2006).
Мощность потока корневых экссудатов в почву заметно варьирует в зависимости от видов деревьев, а также от видового состава микоризных грибов (Liese et al., 2018; Kaiser et al., 2019; Sun et al., 2020; Lei et al., 2023). Максимальная скорость продукции внешнего мицелия в лесной зоне наблюдается в начале лета и в августе (Björk, Ekblad, 2010). До сих пор авторы цитируемых выше работ не дают чёткого ответа на вопрос: перехватывает ли ЭМ из экссудатов весь углерод и весь азот? Так как чехол эктомикоризы имеет плотное сложение (Смит, Рид, 2012), то можно допустить, что полный перехват С и N экссудатов вполне возможен, поскольку эти элементы в органическом веществе являются основными для обеспечения роста биомассы микоризы (роль остальных элементов мы здесь не рассматриваем).
Из полученных от растения-хозяина органических веществ ЭМ тратит до 60% на рост внешнего мицелия (Nehls et al., 2017), что позволяет ей сформировать густую сеть внешнего мицелия и использовать ПОВ всего объёма аккумулятивных горизонтов почвы. В итоге пул внешнего мицелия может доминировать в сообществе почвенной микробиоты: в лесных почвах он составляет 30% от общей биомассы всех микроорганизмов (Högberg, Högberg, 2002). Принимая во внимание, что оставшиеся (после расходов на рост мицелия) 40% углерода экссудатов – это трудно усвояемые органические кислоты, и учитывая реально большие расходы на дыхание микоризы, можно предположить, что ЭМ потребляет органическое вещество всего потока корневых экссудатов.
Экспериментальное определение продуктивности внешнего мицелия представляет собою методологически трудную задачу. Результаты одной из наиболее обстоятельных работ по оценке продуктивности внешнего мицелия ЭМ представлены в таблице. Следует подчеркнуть, что данные этого полевого опыта указывают на достаточно скромные различия продуктивности внешнего мицелия ЭМ в разных лесах, но занижают реальные значения, поскольку в эксперименте (а) для учета мицелия использовался безгумусовый песок, который сильно отличается от окружающей почвы, и (б) игнорировалось “выедание” живого мицелия почвенной микрофауной, его отмирание и последующее потребление другими микроорганизмами. Если сделать поправку на температуру и продолжительность вегетационного периода, то на первом месте по скорости роста внешнего мицелия ЭМ окажутся бореальные леса.
Таблица. Продукция внешнего мицелия ЭМ деревьев по данным полевых экспериментов с прорастанием микоризных грибов в контейнеры с песком в течение вегетационного периода (Ekblad et al., 2013, табл. 1, с. 6-9, пересчитано с сокращениями)
|
Лесные экосистемы (сохранена терминология авторов) |
Количество пробных площадей
|
Биомасса мицелия (кг/га) в конце вегетационного периода |
| Ельники и сосняки бореальных лесов
Ельники бореально-неморальных лесов Ельники и дубняки неморальных лесов |
7
19 15 |
151±28
188±12 138±9 |
Восполнение нехватки (майнинг) азота. Корневые экссудаты, обеспечивая микоризу углеродом для синтеза биомассы, имеют в своем составе, как правило, недостаточно азота, необходимого для полного использования ЭМ всего углерода экссудатов на рост мицелия. Это следует из того, что отношение C:N экссудатов – в среднем около 40 (Kuzyakov et al., 2007), а биомасса мицелия имеет C:N 10-12 (Chertov et al., 2017). Аминокислоты и углеводы экссудатов быстро и полностью потребляются для синтеза биомассы мицелия и на его дыхание, тогда как для использования безазотистых соединений экссудатов требуется дополнительный азот. С целью компенсации этого дефицита сеть внешнего мицелия ЭМ разлагает ПОВ, но не для получения углерода как источника вещества и энергии, а для добычи недостающего микоризе азота (Chalot, Brun, 1998; Tunlid et al., 2017; Nehls et al., 2017). По современным представлениям, разложение ПОВ происходит в два этапа (Manzoni, Porporato, 2009; Blagodatsky et al., 2010; Huang et al., 2018). На первом этапе экзоферменты, выделяемые микроорганизмами, способствуют расщеплению крупных молекул полимеров до растворимых фрагментов, на втором этапе – это растворённое органическое вещество потребляется микроорганизмами и другой биотой для роста и поддержания жизни. Судя по всему, микориза разлагает ПОВ только на первой стадии деполимеризации. Из полученного РОВ микориза выборочно потребляет преимущественно органическое вещество, содержащее азот (Martin et al., 2016; Stuart, Plett, 2020). Это подтверждается данными о большом количестве производимого микоризой РОВ (Högberg, Högberg, 2002; He et al., 2020), но механизм этого процесса в деталях ещё недостаточно изучен. В этом выборочном разложении устойчивого ПОВ с последующим потреблением преимущественно азотистых компонентов заключается основное функциональное отличие внешнего мицелия микоризы от гетеротрофных грибов (Talbot et al., 2013; Choreno-Parra, Treseder, 2024).
Использование аминокислот и углеводов корневых экссудатов на рост мицелия приводит к формированию в почве кислой среды за счёт оставшихся в корневых экссудатах низкомолекулярных карбоновых кислот (Arvieu et al., 2003; Van Hees, 2005). В работах микробиологов по деполимеризации ПОВ в результате его разложения почвенными микроорганизмами это обстоятельство никак не принимается во внимание, а рассматривается только активность экзоферментов микроорганизмов. Однако, кислые экссудаты, безусловно, способствуют гидролизу ПОВ, частично выполняя роль экзоферментов. Достаточно вспомнить прежние методы физико-химического фракционирования ПОВ, применявшиеся в почвоведении в ХХ веке (Пономарёва, 1964).
“Добыча“ или майнинг (mining) азота ЭМ из ПОВ для восполнения его нехватки для роста мицелия аналогичны такому же процессу при ризосферном прайминге ПОВ (Blagodatskaya, Kuzyakov, 2008; Chertov et al., 2022). На майнинг азота ЭМ расходует углерод экссудатов (которые идут на синтез ферментов и дыхание), поэтому при исчерпании углерода экссудатов в микоризном грибе майнинг азота прекращается. По данным (Nehls et al., 2017), микоризные грибы восполняют дефицит азота только для обеспечения собственного роста (в основном внешнего мицелия), что означает отсутствие прямого возврата добытого азота растению-хозяину, как это ранее объяснялось в устоявшихся “гуманных“ представлениях о взаимном обмене ресурсами в симбиозе растений и микоризных грибов [см. исторический очерк С. Смит, Д. Рид (2012), а также (van der Heijen, Horton, 2009)].
Возникает вопрос: как осуществляется поток в корни растения-хозяина углерода и азота c положительным эффектом ЭМ на рост древесных растений?
Опад внешнего мицелия. Густая сеть внешнего мицелия представляет собою быстро обновляющийся компонент биомассы ЭМ с продолжительностью жизни от нескольких дней до нескольких недель (Lukac et al., 2003; Staddon et al., 2003; Smith, Read, 2008; Gorka et al., 2019). Причиной этого может быть, как генетически обусловленный небольшой предельный возраст внешнего мицелия, так и его потребление другими микроорганизмами или почвенной микро- и мезофауной; последнее представляется более вероятным.
Логическим следствием интенсивного оборота внешнего мицелия ЭМ является внутрипочвенный поток специфического легкоразлагаемого опада, который раньше не рассматривался в работах по динамике ПОВ. Поступление этого опада в почву оказалось очень большим, с массой 62% от всего наземного и внутрипочвенного опада (Godbold et al., 2006; Brunner et al., 2013; Ekblad et al., 2013), что позволяет считать его доминирующим в структуре опада лесных сообществ. С позиций круговорота углерода, данный факт существенно меняет общую картину процессов трансформации и разложения опада в лесных и других почвах, и в целом в наземных экосистемах. Можно сказать, что микориза оказалась мощным драйвером динамики ПОВ (Talbot et al., 2008; Hawkins et al., 2023).
Разложение внешнего мицелия. В настоящее время разложение опада внешнего мицелия и чехла ЭМ в почве рассматривается микологами как микробиологический процесс, осуществляемый сапротрофными бактериями и грибами (Tunlid et al., 2017; Martin, 2017; Hanajima et al., 2019), а роль остальной почвенной биоты оценивается как вспомогательная (Jentschke et al., 2013). Это довольно странно, поскольку в экологии и почвенной зоологии уже давно разработана, используется и совершенствуется теория и методология трофических сетей (food web), показывающая сложную структурную организацию процесса потребления организмами жизненных ресурсов в почвах, экосистемах и в целом во всей биосфере (Одум, 1986; de Vries, Caruso, 2016; Geisen et al., 2016; Pausch et al., 2016; Komarov et al., 2017; Chertov et al., 2022).
В почве трофическая сеть разложения органического вещества упрощённо может быть представлена как: “растительный опад → микроорганизмы → микро-, мезо- и макрофауна“. Первые два звена – рассматриваются в рамках микробиологического подхода, согласно которому, основным и, скорее всего, побочным продуктом при разложении органического вещества является РОВ, участвующее далее в формировании ПОВ за счёт роста биомассы и отмирания микроорганизмов (Wang et al., 2021), а также закрепления РОВ на поверхности почвенных минералов (Klink et al., 2022). Механизм закрепления работает, но у него есть серьёзное ограничение по ёмкости фиксации минеральной матрицы и почвенного поглощающего комплекса, что, по-видимому, ограничивает масштабы этого процесса.
Рассмотрение всей пищевой сети, когда базовый уровень микроорганизмов ”выедается” микрофауной простейших и членистоногих (microbial grazers) полностью до 9 раз за год (de Vries et al., 2013) без формирования РОВ, предполагает поступление в почву двух специфических продуктов жизнедеятельности почвенной микрофауны. Это твердофазные экскреты (микрокопролиты) и растворимый аммоний, доступный для корневого потребления растений (Schröter et al., 2003; Holtkamp et al., 2011). На более высоких уровнях пищевой сети (с участием мезо- и макрофауны) тоже всегда образуются копролиты, которые пополняют ПОВ в качестве лабильного и богатого азотом пула. В этом заключается главное различие между двумя подходами (микробиологическим и почвенных пищевых сетей): ограниченные возможности формирования ПОВ при фиксации растворенного органического вещества на поверхностях минералов и защемления в микроагрегатах, а с другой стороны, не лимитированный потенциал секвестрации ПОВ из экскретов почвенной фауны. Наличие связи в системе “устойчивое ПОВ → микоризный гриб → почвенная фауна → лабильное ПОВ (экскреты)” подтверждается присутствием “старого” углерода устойчивого органического вещества почв с возрастом более 1000 лет во фракциях “молодого” лабильного ПОВ (Kalinina et al., 2019).
Выше приведены сведения о том, что внешний мицелий ЭМ добывает азот из ПОВ только для собственных нужд, не “передавая” его растению-хозяину. Если это так, то источником дополнительного азота для растений “от микоризы” может быть только пул лабильного органического вещества экскрементов, образующихся после потребления внешнего мицелия почвенной фауной. Этот пул характеризуется достаточно высокой концентрацией азота (низким отношением C:N), ещё не связан с полуторными окислами и основаниями и не ассоциирован с минералами. Безусловно, это органическое вещество пополняет запасы всего ПОВ, но оно также интенсивно используется почвенными микроорганизмами, что ведет к его разложению и образованию различных форм соединений азота, доступных для растения-хозяина. Таковыми являются не только аммоний и нитраты, но прежде всего аминокислоты. Известно, что доступный для деревьев азот лесных почв на 80% представлен аминокислотами, 5-15% аммонием и 5-11% нитратами (Pena, 2017). Именно этот путь поступления азота отмершего или “съеденного” внешнего мицелия в корни, по-видимому, и обеспечивает эффект улучшения азотного питания древесных растений в результате деятельности ЭМ.
При рассмотрении этого достаточно сложного пути образования доступного для растений азота возникает вопрос о его поступлении в корни: непосредственно в корень или через внешний мицелий и чехол гриба. В эктомикоризном симбиозе вопрос о непосредственном поступлении азота в корень отпадает из-за наличия плотного мицелиального чехла на корневых окончаниях. Тогда почему ЭМ не использует доступный азот также для своего роста? Это может происходить по следующим причинам. Во-первых, потому что внешний мицелий микоризы играет еще и транспортную роль в водном режиме растений, поддерживая потребление воды и транспирационный поток почвенного раствора в растении (Смит, Рид, 2012; в данной статье не рассматривается). Скорость восходящего транспирационного потока почвенного раствора с элементами питания во внешнем мицелии и корнях на порядки выше скорости обратного потока экссудатов, что легко вычисляется по суточной массе воды в транспирационном потоке и экссудации. Эти процессы находятся в противофазе чередования транспирации и экссудации. Можно считать, что высокая скорость потока затрудняет перехват элементов питания из почвенного раствора как внешним мицелием, так и чехлом эктомикоризы. Кроме того, нужно принимать во внимание большое различие концентраций элементов в почвенном растворе и в активной зоне майнинга азота микоризного гриба.
Во-вторых, транзит доступного азота через микоризу возможен потому, что микоризные грибы являются облигатными автохтонными организмами-биотрофами со своими специфическими механизмами «добычи» и особенно ассимиляции азота, используя ансамбль фенолоксидаз. Эти механизмы могут быть не активны в отношении продуктов микробиологического разложения “молодого“ ПОВ. Но, как обсуждалось выше, в составе доступного корням азота содержится до 20% минеральных форм азота. По-видимому, только у нитратов и аммония есть шансы быть ассимилированными во внешнем мицелии и чехле ЭМ. Безусловно, всё это требует экспериментального подтверждения.
Растворение минералов. ЭМ обладает еще одним свойством, которое оказалось неожиданным для почвоведов и биологов. Было обнаружено, что гифы микоризы способны растворять почвенные минералы, оставляя хорошо видимый под микроскопом след на их поверхности (van Breemen et al., 2000; Sun et al., 2019). Растворение минералов – это еще одна функция кислого РОВ, образующегося при майнинге азота ЭМ, о чем упоминалось выше. Данный процесс позволяет понять роль ЭМ в снабжении растений фосфором, кальцием и другими минеральными элементами. Изучение этого процесса развивается, и уже идёт речь о создании научного направления – геомикологии (Gadd, 2007; Smits, Wallander, 2017).
Круг замкнулся: то, что в середине ХХ в. рассматривалось с позиций генезиса лесных почв таёжной зоны и не привлекало особого внимания, а именно, характерная для этих почв кислая среда (преобладание фульвокислот (Пономарева, 1964)), в настоящее время фигурирует под именем “растворимое органическое вещество” как драйвер динамики ПОВ, растворения и трансформации минералов в лесных почвах.
Общая концепция и перспективы. Вышеизложенное позволяет составить общее представление о «работе» ЭМ в отношении динамики ПОВ и её роли в питании древесных растений на основе той информации, которая была в распоряжении авторов. На упрощённой схеме (рисунок) приведены основные пулы органического вещества и “связывающие” их процессы. Для облегчения визуального восприятия, нами не показаны потоки СО2, связанные с дыханием ЭМ при её росте и майнинге азота, а также дыхание микроорганизмов и почвенной фауны. Также не отражены кислые экссудаты и минеральная матрица.
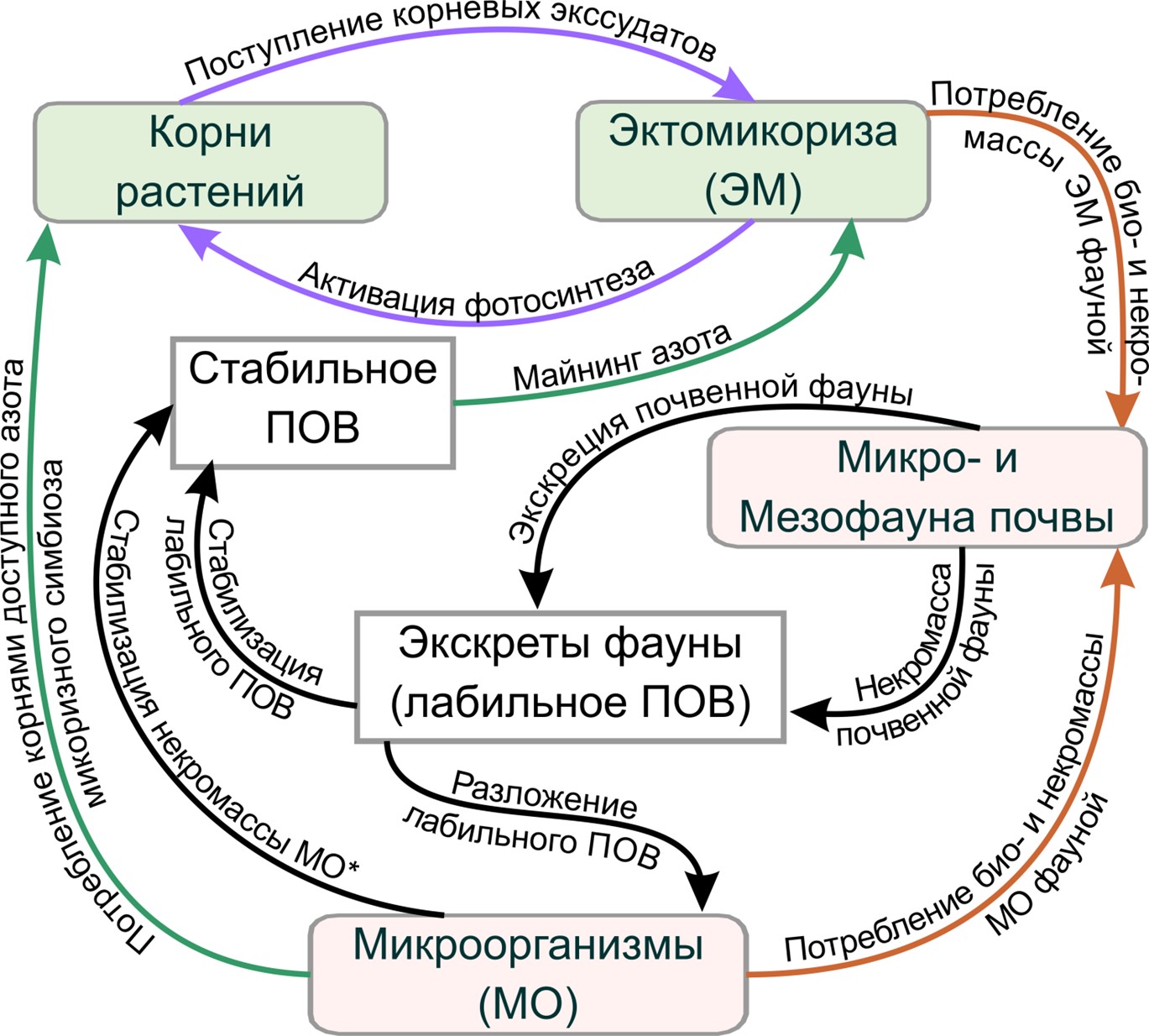
Рисунок 1. Обобщённая концептуальная схема потоков и пулов органического вещества при функционировании эктомикоризы, её вклада в пополнение ПОВ и снабжение растений азотом. Наименования относятся как к пулам, так и к потокам, стрелки только к потокам. На схеме не представлены потоки СО2 при дыхании биоты, растворимое органическое вещество (РОВ), растворение минералов кислыми экссудатами и не отражено поступление доступного азота в корни через внешний мицелий и чехол эктомикоризного гриба с транспирационным потоком воды из почвы.
Рассмотренные данные по функционированию ЭМ с точки зрения цикла углерода и азота в лесных почвах выявили сложную структуру состава образующегося РОВ, в которое входят: а) возможный остаток кислых корневых экссудатов; б) растворённые фрагменты молекул устойчивого ПОВ после деполимеризации для майнинга азота; в) вероятно и РОВ, образующееся при минерализации лабильного органического вещества экскретов почвенной микрофауны. Поэтому состав РОВ потенциально может отражать состояние системы микоризного симбиоза, например, по количеству карбоновых кислот корневых экссудатов в его составе.
Микоризный симбиоз является более сложным и масштабным явлением, чем хорошо изученный ризосферный прайминг. Более того, взаимоотношения микоризы и микробиоты, участвующей в ризосферном прайминге неоднозначны – от взаимовыгодных до конкурентных (Chertov et al., 2022). Конкуренция за один и тот же ресурс представляется наиболее вероятной формой взаимодействия микоризного симбиоза и ризосферного прайминга. Так, обнаружено, что в присутствии арбускулярной микоризы интенсивность ризосферного прайминга существенно снижается (Zhou et al., 2020). Надо полагать, что плотный чехол ЭМ с перехватом почти всех корневых экссудатов даёт мало шансов развития ризосферному праймингу для эффективного функционирования в лесной почве.
За полтора века исследований по микоризной тематике накоплено огромное количество экспериментальных данных по распространению, биологическому разнообразию, строению, функционированию и взаимосвязи микоризы разных типов с другими организмами (Смит, Рид, 2012), Обобщение этих данных требует большой и кропотливой работы. Одним из эффективных методов синтеза научных результатов является математическое моделирование в биологии, которое позволяет в строгой математической форме систематизировать данные о структуре и функционировании организмов, их взаимосвязей, а также функционирования экосистем разного иерархического уровня вплоть до биосферы Земли с целью обоснованного прогнозирования и управления биологическими процессами в современной быстро меняющейся окружающей среде.
Это справедливо и для синтеза данных по микоризе. Появились работы по концептуализации и выработке принципов моделирования такого сложного явления как микоризный синтез (Brunner et al., 2013; Deckmyn et al., 2014) с акцентом на роль микоризы в питании растений. Микоризу начали включать в структуру моделей динамики органического вещества почв, наземных экосистем и в сложные модельные системы как потребителя экссудатов (Meyer et al., 2012). Появились модели, отражающие разные стороны микробного симбиоза, например пространственная модель распространения микоризы (Simard et al., 2012) и, с другой стороны, достаточно детальная модель расхода углерода на потребление азота с учётом конкурентных отношений (Brzostek et al., 2014). Примером хорошей детально параметризованной модели на основе использования всех доступных экспериментальных данных является модель MYCOFON (Meyer et al., 2010, 2012), которая также может быть интегрирована с моделями лесных экосистем.
ЗАКЛЮЧЕНИЕ
Подводя общий итог изложенным выше материалам по вкладу ЭМ древесных растений в разложение и пополнение ПОВ лесных почв, можно сформулировать следующие положения.
- Растение в микоризном симбиозе снабжает гриб элементами питания, но ЭМ ведёт себя активнее растения. Она инфицирует корни, а с восходящим транспирационным потоком влаги направляет в листья вещества, которые активизируют фотосинтез и приводят к возрастанию НБП растения. В результате поток корневых экссудатов увеличивается, что благоприятно сказывается на росте микоризы.
- ЭМ потребляет на собственный рост (прежде всего, внешнего мицелия) весь азот корневых экссудатов, поступающих в ЭМ из растения-хозяина.
- Разложение ПОВ микоризой осуществляется для восполнения нехватки азота на рост внешнего мицелия. Таким образом, это избирательный майнинг азота из ПОВ с повышенным содержанием азота. Фактически, микоризный гриб “работает только на себя”, поэтому вклад ЭМ в разложение ПОВ невелик, что отличает микоризные грибы от всех сапротрофных микроорганизмов.
- Внешний мицелий ЭМ имеет короткий срок жизни (от нескольких дней до нескольких недель), что определяет высокую скорость оборота его биомассы в почве. Соответственно, опад внешнего мицелия можно считать доминирующим в структуре общего опада в лесных экосистемах. Это определяет значимую роль ЭМ в динамике ПОВ лесных почв.
- Разложение опада ЭМ происходит с активным участием почвенной микро- и мезофауны разных трофических уровней пищевых сетей, что обеспечивает пополнение ПОВ за счёт твердофазных продуктов метаболизма почвенной фауны (лабильного органического вещества экскретов). При чисто микробиологическом разложении, где побочным продуктом может быть только РОВ, физико-химическое закрепление органического вещества на почвенных минералах ограничено ёмкостью поглощающего комплекса почвы.
- Дополнительный азот для потребления растением-хозяином в микоризном симбиозе образуется за счёт микоризного майнинга устойчивого ПОВ, но в этом случае азот становится доступным для корней только после разложения микроорганизмами части лабильного органического вещества экскретов.
- “Остаточные” корневые экссудаты, которые после “выедания” ЭМ углеводов и аминокислот для своего роста имеют более высокую кислотность и способны растворять почвенные минералы, обеспечивают доступ почвенной биоты и корней растений к фосфору и другим элементам.
- Положительное влияние ЭМ на ПОВ связано с улучшением корневого питания и роста древесных растений, что приводит к общему увеличению опада в лесных сообществах и, соответственно, к косвенному вкладу ЭМ в пополнение ПОВ лесных почв.
С точки зрения структуры наземных и, особенно, лесных экосистем можно заключить, что микоризный симбиоз оказывается не диполем – «растение + микориза», как это укоренилось в научных представлениях, а триадой – «растение + микориза + почвенная биота» с мощным воздействием на оптимизацию эдафической среды наземных экосистем. Без третьего компонента — почвенной биоты — положительный эффект микоризы на рост растений просто не сможет проявиться из-за «эгоизма» гриба. Корни растений являются каркасом и стартёром системы микоризного симбиоза, сама микориза служит драйвером процессов, а остальная почвенная биота осуществляет выход продуктов симбиоза в почву и обратно в корни растений. Высказанные положения основаны на анализе доступных авторам материалов, некоторые из этих положений могут показаться спорными. Несомненно, необходимы дальнейшие исследования, как по широкому спектру проблем взаимодействия растения и ЭМ, так и в рамках положения о триаде “растение — микориза — почвенная биота” в микоризном симбиозе.
БЛАГОДАРНОСТИ
Работа выполнена в рамках реализации важнейшего инновационного проекта государственного значения «Разработка системы наземного и дистанционного мониторинга пулов углерода и потоков парниковых газов на территории Российской Федерации, обеспечение создания системы учёта данных о потоках климатически активных веществ и бюджете углерода в лесах и других наземных экологических системах» (рег. № 123030300031) и государственного задания ФИЦ ПНЦБИ (рег. № 122040500037-6).
СПИСОК ЛИТЕРАТУРЫ
Аккумуляция углерода в лесных почвах и сукцессионный статус лесов. Отв. Ред. Лукина Н. В. М.: КМК, 2018. 232 с.
Базилевич Н. И. Биотический круговорот на пяти континентах: азот и зольные элементы в природных наземных экосистемах. Новосибирск: Изд-во СО РАН. 2008. 376 с.
Вацке Р. Использование микоризы в садоводстве как метод решения проблем в области экологического возделывания растений // Плодоводство и ягодоводство России. 2005. Т. 14. С. 234–244.
Весёлкин Д. В. Морфологическая изменчивость и адаптивное значение эктомикориз хвойных (Pinaceae Lindl.). Автореф. дисс. докт. биол. наук. Екатеринбург, 2013. 40 с.
Весёлкин Д. В. Соотношение объемов гриба и древесных тканей в эктомикоризных корнях хвойных // Лесоведение. 2015. № 2. С. 140–146.
Весёлкин Д. В., Лукина Н. В., Чибрик Т. С. Соотношение микоризных и немикоризных видов растений в первичных техногенных сукцессиях // Экология. 2015. № 5. С. 345–352.
Волобуева О. Г. Симбиотическая азотфиксация как фактор экологической безопасности и плодородия почвы // Вестник РУДН, сер. Экология и безопасность жизнедеятельности. 2011. № 1. С. 53–60.
Воронина Е. Ю. Микоризы и их роль в формировании сообществ // Вестник Московского университета. Серия 16: Биология. 2006. № 4. С. 17–27.
Воронина Е. Ю. Микоризы в наземных экосистемах: экологические, физиологические и молекулярно-генетические аспекты микоризных симбиозов // Микология сегодня. Ред. Ю.Т. Дьяков, А.Ю. Сергеев. Национальная Академия Микологии. М. 2007. С. 101–192.
Дудка В. А., Малышева Е. Ф., Малышева В. Ф. Жукова Е. А. Микориза липы (Tilia spp.) в искусственных насаждениях Санкт-Петербурга // Микология и фитопатология. 2021. Т. 55. № 1. С. 11–35.
Иозус А. П., Завьялов А. А., Бойко С. Ю. Особенности влияния микоризы на приживаемость и биохимический состав сеянцев сосны обыкновенной в сухой степи Нижнего Поволжья // Успехи современного естествознания. 2019. № 6. С. 23–27.
Колмаков П. Ю., Кисова А. С. Разнообразие эктомикориз Picea abies в естественных местообитаниях Белорусского Поозерья // Материалы II Международной конференции, 20-23 сентября 2016 г. „Биология, систематика и экология грибов и лишайников в природных экосистемах и агрофитоценозах“. Минск, 2016. С. 123–125.
Лавренов Н. Г., Зернов А. С., Кипкеев А. М., Текеев Д. К., Семенова Р. Б., Ахметжанова А. А., Переведенцева Л. Г., Судзиловская Н. А., Корнеечева М. Ю., Онипченко В.Г. Микориза растений в экстремальных условиях: альпийские ковры Армении // Журнал общей биологии. 2017. Т. 78. № 4. С. 80–85.
Лукина Н. В., Чибрик Т. С., Глазырина М. А., Филимонова Е. И. Динамика восстановления растительности и микоризы на рекультивированных и нерекультивированных участках золоотвала Верхнетагильской ГРЭС (Средний Урал) // Экосистемы. 2019. № 20 (50). С. 188–196.
Макаров М. И. Роль микоризы в трансформации соединений азота в почве и в азотном питании растений (обзор) // Почвоведение. 2019. № 2. С. 220–233.
Одум Ю. Экология. М.: Мир, 1986. Т.1. 328 с.; Т.2. 376 с.
Пономарева В. В. Теория подзолообразовательного процесса. Биохимические аспекты. М.: Наука. 1964. 379 с.
Сергеева М. Н. Грибы. Большая энциклопедия. М.: Изд. Аст. 2022. 256 с.
Смит С. Э., Рид Д. Дж. Микоризный симбиоз. М.: КМК. 2012. 776 с.
Титлянова А. А., Самбуу А. Д. Сукцессии в травяных экосистемах. Новосибирск: Изд-во СО РАН. 2016. 191 с.
Тиунов А. В. Трофические сети в почве бореального леса: в поисках значения микоризы // Проблемы почвенной зоологии. Материалы XVII Всероссийского совещания по почвенной зоологии, посвященного 75-летию со дня рождения члена-корреспондента РАН Д. А. Криволуцкого / Под ред. Б. Р. Стригановой. 2014. С. 220–222.
Хофрихтер Р. Грибы. Обитатели скрытого мира. М.: Изд. Азбука. 2021. 280 с.
Aasfar A., Aasfar A., Bargaz A., Yaakoubi K., Hilali A., Bennis I., Zeroual Y., Meftah Kadmiri I. Nitrogen Fixing Azotobacter Species as Potential Soil Biological Enhancers for Crop Nutrition and Yield Stability // Frontiers in microbiology. 2021. Vol. 12. Article 628379.
Agerer R., Hartmann A., Pritsch K., Raidl S., Schloter M., Verma R., Weigt R. Plants and Their Ectomycorrhizosphere: Cost and Benefit of Symbiotic Soil Organisms // Growth and Defence in Plants. Resource Allocation at Multiple Scales. 2012. Vol. 220. P. 213–242.
Akhmetzhanova A. A., Soudzilovskaia N. A., Onipchenko V. G., Cornwell W. K., Agafonov V. A., Selivanov I. A., Cornelissen J. H. C. A rediscovered treasure: Mycorrhizal intensity database for 3000 vascular plant species across the former Soviet Union // Ecology. 2012. Vol. 93. P. 689–690.
Albornoz F. E., Dixon K. W. Lambers H. Revisiting mycorrhizal dogmas: Are mycorrhizas really functioning as they are widely believed to do? // Soil Ecology Letters. 2021. Vol. 3. P. 73–82.
Arvieu J. C., Leprince F., Plassard C. Release of oxalate and protons by ectomycorrhizal fungi in response to P-deficiency and calcium carbonate in nutrient solution // Annals of Forest Science. 2003. Vol. 60. P. 815–821.
Averill C., Bhatnagar J. M., Dietze M. C., Pearse W. D., Kivlin S. N. Global imprint of mycorrhizal fungi on whole-plant nutrient economics // PNAS. 2019. Vol. 116. No. 46. P. 23163–23168.
Bastida F., García C., Fierer N., Eldridge D. J., Bowker M. A., …, & Delgado-Baquerizo M. Global ecological predictors of the soil priming effect // Nature Communications. 2019. Vol. 10. P. 3481.
Becquer A., Guerrero C., Janice G., Eibensteiner L., Houdinet G., Bücking H., Zimmermann S. D., Garcia K. The ectomycorrhizal contribution to tree nutrition // Advances in Botanical Research. 2019. Vol. 89. P. 77–126.
Betekhtina А. А., Veselkin D. V. Mycorrhizal and non-mycorrhizal dicotyledonous herbaceous plants differ in root anatomy: evidence from the Middle Urals, Russia // Symbiosis. 2019. Vol. 77. No. 2. P. 133–140.
Björk R. G., Ekblad A. Extramatrical mycelia production and turnover in two drained Norway spruce forests / I. Brunner (Ed.) Belowground carbon turnover in European forests – State of the art. COST Action FP0803 Conference, January 26–28, 2010. Swiss Federal Institute for Forest, Snow and Landscape Research WSL 2010. Birmensdorf, 2010. P. 631–633.
Blagodatskaya Е., Kuzyakov Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review // Biology and Fertility of Soils. 2008. Vol. 45. P. 115–131.
Blagodatsky S., Blagodatskaya E., Yuyukina T., Kuzyakov Y. Model of apparent and real priming effects: Linking microbial activity with soil organic matter decomposition // Soil biology and biochemistry. 2010. Vol. 42. Р. 1275–1283.
Bönisch E., Blagodatskaya E. Dirozo R., Ferlian O., Fichtner A., …, & Eisenhauer N. Mycorrhizal type and tree diversity affect foliar elemental pools and stoichiometry // The New Phytologist. 2024. PMID: 38594212. 15 p.
Brundrett M. C., Tedersoo L., Evolutionary history of mycorrhizal symbioses and global host plant diversity // The New Phytologist. 2008. Vol. 220. P. 1108–1115.
Brundrett M. S. Diversity and classification of mycorrhizal associations // Biological reviews. 2004. Vol. 79. P. 473–495.
Brunner I., Børja I., Dalsgaard L., Deckmyn G., Ekblad A., …, & Vanguelova E. Belowground carbon turnover of European forests: fine roots, mycorrhizal mycelia, soil organic matter and soil models. A Technical Report for National C reporters, LULUCF experts and ecosystem modellers. COST Action FP0803. Birmensdorf, Swiss Federal Institute for Forest, Snow and Landscape Research WSL. 2013. 67 p.
Brzostek E. R., Fishe J. B., Phillip R. P. Modeling the carbon cost of plant nitrogen acquisition: Mycorrhizal trade-offs and multipath resistance uptake improve predictions of retranslocation // Journal of Geophysical Research: Biogeosciences. 2014. Vol. 119 (8). P. 1684–1697.
Canarini A., Kaiser C., Merchant A., Richter A., Wanek W. Root exudation of primary metabolites: Mechanisms and their roles in plant responses to environmental stimuli // Frontiers in Plant Science. 2019. Vol. 10. Р. 157–165.
Chalot M., Brun A. Physiology of organic nitrogen acquisition by ectomycorrhizal fungi and ectomycorrhizas // FEMS Microbiology Reviews. 1998. Vol. 22. P. 21–44.
Chertov O., Komarov A., Shaw C., Bykhovets S., Frolov P., …, & Shashkov M. Romul_Hum — A model of soil organic matter formation coupling with soil biota activity. II. Parameterisation of the soil food web biota activity // Ecological Modelling. 2017. Vol. 345. P. 125–139.
Chertov O., Kuzyakov Y., Priputina I., Frolov P., Shanin V., Grabarnik P. Modelling the rhizosphere priming effect in combination with soil food webs to quantify interaction between living plant, soil biota and soil organic matter // Plants. 2022. Vol. 11. Article 2605.
Choreno-Parra E. M., Treseder K. K. Mycorrhizal fungi modify decomposition: a meta analysis // The New Phytologist. 2024. Vol. 242. No. 6. P. 2763–2774.
de Vries F. T., Caruso T. Eating from the same plate? Revisiting the role of labile carbon inputs in the soil food web // Soil biology and biochemistry. 2016. Vol. 102. P. 4–9.
de Vries F. T., Thébault E., Liiri M., Birkhofer K., Tsiafouli M. A., et al. Soil food web properties explain ecosystem services across European land use systems // Proceedings of the National Academy of Sciences. 2013. Vol. 110. P. 14296–14301.
Deckmyn G., Meyer A., Smits M.M., Ekblad A., Greben T., Komarov A., Kraigher H. Simulating ectomycorrhizal fungi and their role in carbon and nitrogen cycling in forest ecosystems // Canadian Journal of Forest Research. 2014. Vol. 44. No. 6. P. 535–553.
Ekblad A., Wallander H., Godbold D. L., Cruz C., Johnson D., …, & Plassard C. The production and turnover of extramatrical mycelium of ectomycorrhizal fungi in forest soils: role in carbon cycling of extramatrical mycelium (EMM) of mycorrhizal fungi in carbon (C) cycling in ecosystems // Plant Soil. 2013. Vol. 366. P. 1–27.
Fogel R., Hunt G. Contribution of mycorrhizal and soil fungi to nutrient cycling in a Douglas-fir ecosystem // Canadian Journal of Forest Research. 1983. Vol. 13 (2). P. 219–232.
Gadd G. M. Geomycology: biogeochemical transformations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation // Mycological research. 2007. Vol 111. P. 3–49. DOI: 10.1016/j.mycres.2006.12.001.
Geisen S., Koller R., Hünninghaus M., Dumack K., Urich T., Bonkowski M. The soil food web revisited: Diverse and widespread mycophagous soil protists // Soil Biology and Biochemistry. 2016. Vol. 94. P. 10–18. DOI: 10.1016/j.soilbio.2015.11.010.
Godbold D. L., Hoosbeek M. R., Lukac M., Cotrufo M. F., Janssens I. A., …, & Prescott A. Mycorrhizal hyphal turnover as a dominant process for C input into soil organic matter // Plant Soil. 2006. Vol. 281. P.15–24.
Gorka S., Dietrich M., Mayerhofer W., Gabriel R., Wiesenbauer J., …, & Kaiser C. Rapid transfer of plant photosynthates to soil bacteria via ectomycorrhizal hyphae and its interaction with nitrogen availability // Frontiers in Microbiology. 2019. Vol. 10. Article 168. DOI: 10.3389/fmicb.2019.00168.
Guignabert A., Delerue F., Gonzalez M., Augusto L., Bakker M. Effects of management practices and topography on ectomycorrhizal fungi of maritime pine during seedling recruitment // Forests. 2018. Vol. 9. No. 5. DOI: 10.3390/f9050245.
Hanajima D., Aoyagi T., Hori T. Dead bacterial biomass-assimilating bacterial populations in compost revealed by high-sensitivity stable isotope probing // Environment International. 2019. Vol. 133. Article 105235. DOI: 10.1016/j.envint.2019.
Hawkins H.-J., Cargill R. I. M., Van Nuland M. E., Hagen S. C., Field K. J., Sheldrake M., Soudzilovskaia N., Kiers E. T. Review. Mycorrhizal mycelium as a global carbon pool // Current Biology. 2023. Vol. 33. No. 11. Article R560–R573.
He Y. H., Cheng W. X., Zhou L. Y., Shao J. J., Liu H. Y., Zhou H. M., Zhu K., Zhou X. H. Soil DOC release and aggregate disruption mediate rhizosphere priming effect on soil C decomposition // Soil Biology and Biochemistry. 2020. Vol. 144. Article 107787. DOI: 10.1016/j.soilbio.2020.107787.
Hobbie E. A. Carbon allocation to ectomycorrhizal fungi correlates with belowground allocation in culture studies // Ecology. 2006. Vol. 87. P. 563–569. DOI: 10.1890/05-0755.
Hobbie E. A., Ouimette A. P., Schuur E. A. G., Kierstead D., Trappe J. M., Bendiksen K., Ohenoja E. Radiocarbon evidence for the mining of organic nitrogen from soil by mycorrhizal fungi // Biogeochemistry. 2013. Vol. 114. P. 381–389. DOI: 10.1007/s10533-012-9779-z.
Högberg M. N., Högberg P. Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil // The New Phytologist. 2002. Vol. 154. P. 791–795. DOI: 10.1046/j.1469-8137.2002.00417.x.
Holtkamp R., van der Wal A., Kardol P., van der Putten W. H., de Ruiter P. C., Dekker S. C. Modelling C and N mineralisation in soil food webs during secondary succession on ex-arable land // Soil Biology and Biochemistry. 2011. Vol. 43. P. 251–260.
Huang Ye, Guenet B., Ciais P., Janssens I.A., Soong J. L., Wang Y., Goll D., Blagodatskaya E., Huang Yu. ORCHIMIC (v1.0), A microbe-mediated model for soil organic matter decomposition // Geoscientific Model Development. 2018. Vol. 11. P. 2111–2138. DOI: 10.5194/gmd-11-2111-2018.
Jacquemyn H., Merckx V. S. F. T. Mycorrhizal symbioses and the evolution of trophic modes in plants // Journal of Ecology. 2019. Vol. 107. P. 1567–1581. DOI: 10.1111/1365-2745.13165.
Jentschke G., Bonkowski M., Godbold D. L., Scheu S. Soil protozoa and forest tree growth: non-nutritional effects and interaction with mycorrhizae // AGRIS. 2013. Vol. 20. Р. 263–269. DOI: 10.1007/BF00336088.
Johansson E. M., Fransson P. M. A., Finlay R. D., van Hees P. A. W. Quantitative analysis of root and ectomycorrhizal exudates as a response to Pb, Cd and as stress // Plant Soil. 2008. Vol. 313. P. 39–54. DOI: 10.1007/s11104-008-967.
Johnson N. C., Graham J. H. The continuum concept remains a useful framework for studying mycorrhizal functioning // Plant Soil. 2013. Vol. 363. P. 411−419. DOI: 10.1007/s11104-012-1406.
Johnson N. C., Wilson G. W. T., Wilson J. A., Miller R. M., Bowker M. A. Mycorrhizal phenotypes and the law of the minimum // The New Phytologist. 2015. Vol. 205. P. 1473–1484. DOI: 10.1111/nph.13172.
Jones D. L., Hodge A., Kuzyakov Y. Plant and mycorrhizal regulation of rhizodeposition // The New Phytologist. 2004. Vol. 163. P. 459–480. DOI: 10.1111/j.1469-8137.2004.01130.x.
Kaiser C., Merchant A., Richter A., Wanek W. Root exudation of primary metabolites: Mechanisms and their roles in plant responses to environmental stimuli // Frontiers in Plant Science. 2019. Vol. 10. Article 157. DOI: 10.3389/fpls.2019.00157.
Kalinina O., Cherkinsky A., Chertov O., Goryachkin S., Kurganova I., Lopes de Gerenyu V., Lyuri D., Kuzyakov Y., Giani L. Post-agricultural restoration: Implications for dynamics of soil organic matter pools // Catena. 2019. Vol. 181. Article 104096. DOI: 10.1016/j.catena.2019.104096.
Klink S., Keller A. B., Wildae A. J., Baumert V. L., Gube M., …, & Pausch J. Stable isotopes reveal that fungal residues contribute more to mineral-associated organic matter pools than plant residues // Soil Biology and Biochemistry. 2022. Vol. 168. Article 108634. DOI: 10.1016/j.soilbio.2024.109323.
Komarov A., Chertov O., Bykhovets S., Shaw C., Nadporozhskaya M., … & Zubkova E. Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing // Ecological Modelling. 2017. Vol. 345. P. 113–124. DOI: 10.1016/j.ecolmodel.2016.08.007.
Kranabetter J. M. Ectomycorrhizal fungi and the nitrogen economy of conifers—implications for genecology and climate change mitigation // Botany. 2014. Vol. 92. P. 417–423. DOI: 10.1139/cjb-2013-0198.
Kuzyakov Y., Hill P. W., Jones D. L. Root exudate components change litter decomposition in a simulated rhizosphere depending on temperature // Plant Soil. 2007. Vol. 290. P. 293–305.
Lei X., Shen Y., Zhao J., Huang J., Wang H., Yu Y., Xiao C. Review: root exudates mediate the processes of soil organic carboni and efflux // Plants. 2023. Vol. 12 (3). Article 630. DOI: 10.3390/plants12030630.
Liese R., Lübbe T., Albers N. W., Meier I. C. The mycorrhizal type governs root exudation and nitrogen uptake of temperate tree species // Tree Physiology. 2018. Vol. 38. P. 83–95. DOI: 10.1093/treephys/tpx131.
Lindahl B. D., Tunlid A. Ectomycorrhizal fungi – potential organic matter decomposers, yet not saprotrophs // The New Phytologist. 2015. Vol. 205. No. 4. P. 1443–1447.
Lukac M., Calfapietra C., Godbold D. L. Production, turnover and mycorrhizal colonization of root systems of three Populus species grown under elevated CO2 (POPFACE) // Global Change Biology. 2003. Vol. 9. P. 838–848. DOI: 10.1046/j.1365-2486.2003.00582.x.
Manzoni S., Porporat A. Soil carbon and nitrogen mineralization: theory and models across scales // Soil Biology and Biochemistry. 2009. Vol. 41. P. 1355–1379. DOI: 10.1016/j.soilbio.2009.02.031.
Martin F. (Ed.). Molecular Mycorrhizal Symbiosis. John Wiley & Sons, Hoboken, New Jersey, 2017. 506 p.
Martin F., Kohler, A., Murat C., Veneault-Fourrey C., Hibbett D. S. Unearthing the roots of ectomycorrhizal symbioses // Nature Reviews Microbiology. 2016. Vol. 14 (12). P. 760–773. DOI: 10.1038/nrmicro.2016.149
Meyer A., Grote R., Butterbach-Bahl K. Integrating mycorrhiza in a complex model system: effects on ecosystem C and N fluxes // European Journal of Forest Research // 2012. Vol. 131(6). P. 1809–1831. DOI: 10.1007/s10342-012-0634-5.
Meyer A., Grote R., Pol A. Simulating mycorrhiza contribution to forest C- and N cycling — the MYCOFON model // Plant Soil. 2010. Vol. 327. P. 493–517. DOI: 10.1007/s11104-009-0017-y.
Mus F., Crook M. B., Garcia K., Garcia Costas A., Geddes B. A., …, & Peters J. W. Symbiotic nitrogen fixation and the challenges to its extension to nonlegumes // Applied and Environmental Microbiology. 2016. Vol. 82. P. 3698 –3710. DOI: 10.1128/AEM.01055-16.
Nehls U., Das A., Neb D. Carbohydrate metabolism in ectomycorrhizal symbiosis [in:] F. Martin (Ed.) Molecular Mycorrhizal Symbiosis. Chapter 10. John Wiley & Sons, 2017. P. 161–177.
Nehls U., Göhringer F., Wittulsky S., Dietz S. Fungal carbohydrate support in the ectomycorrhizal symbiosis: a review // Plant Biology. 2010. Vol. 12 (2). P. 292–301. DOI: 10.1111/j.1438-8677.2009.00312.x.
O’Hanlon R. Below-ground ectomycorrhizal communities: the effect of small scale spatial and short term temporal variation // Symbiosis. 2012. Vol. 57. P. 57–71. DOI: 10.1007/s13199-012-0179.
Pang W., Zhang P., Zhang Y., Zhang X., Huang Y., Zhang T., Liu B. The ectomycorrhizal fungi and soil bacterial communities of the five typical tree species in the Junzifeng National Nature Reserve, Southeast China // Plants. 2023. Vol. 12. Article 3853. DOI: 10.3390/plants12223853.
Pausch J., Kramer S., Scharroba A., Scheunemann N., Butenschoen O., …, & Ruess L. Small but active – pool size does not matter for carbon incorporation in belowground food webs // Functional Ecology. 2016. Vol. 30. P. 479–489. DOI: 10.1111/1365-2435.12512.
Pausch J., Tian, J., Riederer M., Kuzyakov Y. Estimation of rhizodeposition at field scale: Upscaling of a 14C labeling study // Plant Soil. 2013. Vol. 364. P. 273–285. DOI: 10.1007/s11104-012-1363-8.83.
Pena R. Nitrogen aquisition in ectomicorrhizal symbiosis [in:] Martin F. (Ed.) Molecular Mycorrhizal Symbiosis. Chapter 11. John Wiley & Sons, 2017. P. 178–196.
Rineau F., Shah F., Smits M.M., Persson P., Johansson T., Carleer R., Troein C., Tunlid A. Carbon availability triggers the decomposition of plant litter and assimilation of nitrogen by an ectomycorrhizal fungus // The ISME journal. 2013. Vol. 7 (10). P. 2010–2022. DOI: 10.1038/ismej.2013.91.
Saifuddin M., Bhatnagar J. M., Phillips R. P., Finzi A. C. Ectomycorrhizal fungi are associated with reduced nitrogen cycling rates in temperate forest soils without corresponding trends in bacterial functional groups // Oecologia. 2021. Vol. 196. P. 863–875.
Savolainen T., Kytöviita M.-M. Mycorrhizal symbiosis changes host nitrogen source use // Plant Soil. 2022. Vol. 471. P. 643–654.
Schröter D., Wolters V., De Ruiter P. C and N mineralisation in the decomposer food webs of a European forest transect // Oikos. 2003. Vol. 102. P. 294–308. DOI: 10.1034/j.1600-0579.2003.12064.x.
Simard S. W., Beiler K. J., Bingham M. A., Deslippe J. R., Philip L. J., Teste F. P. Mycorrhizal networks: mechanisms, ecology and modelling // Fungal Biology Reviews. 2012. Vol. 26. P. 39–60. DOI: 10.1016/j.fbr.2012.01.001.
Smith S. E., Read D. J. Mycorrhizal symbiosis. 3rd ed. Academic Press, Amsterdam, 2008. DOI: 10.1016/B978-0-12-370526-6.X5001-6.
Smits M., Wallander H. Role of Mycorrhizal Symbiosis in Mineral Weathering and Nutrient Mining from Soil Parent Material. Chapter 3 // N. C. Johnson, C. Gehring, J. Jansa (Eds.) Mycorrhizal Mediation of Soil. Elsevier, 2017. P. 35–46. DOI: 10.1016/B978-0-12-804312-7.00003-6.
Soudzilovskaia N. A., Vaessen S., van’t Zelfde M., Raes N. Global patterns of mycorrhizal distribution and their environmental drivers [in:] L. Tedersoo (Ed.) Biogeography of Mycorrhizal Symbiosis. Ecological Studies. Vol. 230. Springer, 2017. P. 223–235. DOI: 10.1007/978-3-319-56363-3_11.
Staddon P. L. Mycorrhizal fungi and environmental change: the need for a mycocentric approach // The New Phytologist. 2005. Vol. 167. P. 635–637.
Staddon P. L., Ramsey C. B., Ostle N., Ineson P., Fitter A. H. Rapid turnover of hyphae of mycorrhizal fungi determined by AMS microanalysis of 14C // Science. 2003. Vol. 300. P. 1138–1140. DOI: 10.1126/science.1084269.
Stuart E. K., Plett K. L. Digging deeper: in search of the mechanisms of carbon and nitrogen exchange in ectomycorrhizal symbioses // Frontiers in Plant Science. 2020. Vol. 10. Article 1658. DOI: 10.3389/fpls.2019.01658.
Sun L., Ataka M., Han M., Han Y., Gan D., Xu T., Guo Y., Zhu B. Root exudation as a major competitive fine-root functional trait of 18 coexisting species in a subtropical forest // The New Phytologist. 2020. Vol. 229 (1). P. 259–271. DOI: 10.1111/nph.16865.
Sun Q., Fu Z., Finlay R., Lian B. Transcriptome analysis provides novel insights into the capacity of the ectomycorrhizal fungus Amanita pantherina to weather K-containing feldspar and apatite // Applied and Environmental Microbiology. 2019. Vol. 85 (15). Article e00719-19.
Talbot J. M., Allison S. D., Treseder K. K. Decomposers in disguise: Mycorrhizal fungi as regulators of soil C dynamics in ecosystems under global change // Functional Ecology. 2008. Vol. 22. P. 955–963. DOI: 10.1111/j.1365-2435.2008.01402.x.
Talbot J. M., Bruns T. D., Smith D. P., Branco S., Glassman S. I., Erlandson S., Vilgalys R., Peay K. G. Independent roles of ectomycorrhizal and saprotrophic communities in soil organic matter decomposition // Soil Biology and Biochemistry. 2013. Vol. 57. P. 282–291. DOI: 10.1016/j.soilbio.2012.10.004.
Tedersoo L. (Ed.) Biogeography of Mycorrhizal Symbiosis. Vol. 230. Springer, 2017. 566 p. DOI: 10.1007/978-3-319-56363-3.
Tedersoo L., Bahram M., Zobel M. How mycorrhizal associations drive plant population and community biology // Science. 2020. Vol. 367 (6480). Article eaba1223. DOI: 10.1126/science.aba1223.
Threatt S. D., Rees D. C. Biological nitrogen fixation // Theory, Practice, and Reality: A Perspective on the Molybdenum Nitrogenase System // FEBS Letters. 2023. Vol. 597(1). P. 45–58. DOI: 10.1002/1873-3468.14534.
Tunlid A., Floudas D., Koide R., Rineau F. Soil organic matter decomposition mechanisms in ectomycorrhizal fungi, molecular // Mycorrhizal Symbiosis. 2017. 10.1002/9781118951446. P. 257–275. DOI: 10.1002/9781118951446.
van Breemen N., Finlay R., Lundström U., Jongmans A. G., Giesler R., Olsson M. Mycorrhizal weathering: A true case of mineral plant nutrition? // Biogeochemistry. 2000. Vol. 49. P. 53–67. DOI: 10.1023/A:1006256231670.
Van der Heijden M. G. A., Horton T. R. Socialism in Soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems // Journal of Ecology. 2009. Vol. 97. P. 1139–1150. DOI: 10.1111/j.1365-2745.2009.01570.x.
Van Hees P. A. W. Oxalate and ferricrocin exudation by the extramatrical mycelium of an ectomycorrhizal fungus in symbiosis with Pinus sylvestris // The New Phytologist. 2005. Vol. 169 (2). P. 367–378. DOI: 10.1111/j.1469-8137.2005.01600.x.
Wallander H., Nilsson L. O., Hagerberg D., Bååth E. Estimation of the biomass and seasonal growth of external mycelium of ectomycorrhizal fungi in the field // The New Phytologist. 2001. Vol. 151. P. 753–760. DOI: 10.1046/j.0028-646x.2001.00199.x.
Wang B., An S., Liang C., Liu Y., Kuzyakov Y. Microbial necromass as the source of soil organic carbon in global ecosystems // Soil Biology and Biochemistry. 2021. Vol. 162. Article 108422. DOI: 10.1016/j.soilbio.2021.108422.
Wang Q., Xing Y., Luo Xi, Liu G., Yan G. Changes in ectomycorrhizal and saprophytic fungal communities during a secondary succession of temperate forests // European Journal of Forest Research. 2023. Vol. 142. P. 1313–1323.
Zhou J., Zang H., Loeppmann S., Gube M., Kuzyakov Y., Pausch J. Arbuscular mycorrhiza enhances rhizodeposition and reduces the rhizosphere priming effect on the decomposition of soil organic matter // Soil Biology and Biochemistry. 2020 Vol. 140. Article 107641. DOI: 10.1016/j.soilbio.2019.107641.
Рецензент: д. б. н. Крышень А. М.





