- DOI: 10.31509/2658-607x-202474-157
- УДК 581.55
Годичный прирост мхов в среднетаежных сосняках кустарничковых Республики Карелия
![]()
© 2024 Н. В. Геникова1*, А. Ю. Карпечко1, Р. П. Обабко1, В. Л. Миронов2,
С. А. Кутенков2
1Институт леса КарНЦ РАН, Россия, Республика Карелия, 185910, Петрозаводск, ул. Пушкинская, 11
2Институт биологии КарНЦ РАН, Россия, Республика Карелия, 185910, Петрозаводск, ул. Пушкинская, 11
*E-mail: genikova@krc.karelia.ru
Поступила в редакцию: 11.10.2024
После рецензирования: 11.11.2024
Принята к печати: 26.11.2024
В статье приводятся морфологические признаки определения годичного прироста для распространенных видов зеленых и сфагновых мхов, произрастающих в Карелии. Рассмотрены Pleurozium schreberi, Hylocomium splendens, мхи р. Dicranum, Aulacomium palustre, Ptilium crista—castrensis, Polytrichum commune, мхи р. Sphagnum. Отличительные признаки годичных приростов выявлены на основе литературных данных и в ходе собственных наблюдений. Для апробации методики проведен анализ влияния разных типов сосновых лесов и кроны сосны на значения годичного прироста Pleurozium schreberi, Hylocomium splendens и Dicranum polysetum. Выявлено, что основной вклад в суммарную годичную продукцию мохового покрова обследованных сосновых лесов вносит Pleurozium schreberi (41.4±6.3 г/м2). Средние значения прироста текущего года для Hylocomium splendens и Dicranum polysetum составляют 5.9±1.1 и 4.8±1.1 г/м2 соответственно. В сосняках черничных на песчаных почвах годичная продукция Pleurozium schreberi под кроной сосны в два раза выше по сравнению с межкроновым пространством (61.2±11.8 и 29.9±9.3 г/м2 соответственно).
Ключевые слова: методика, годичная продукция, фитомасса, зеленые мхи, сфагновые мхи, бореальные леса
Растения живого напочвенного покрова – важный компонент лесных фитоценозов, обеспечивающий устойчивость экосистем (Костина и др., 2016; Телеснина и др., 2024; Cornelissen et al., 2007). В лесных сообществах таежной зоны мхи являются одним из основных представителей напочвенной растительности. Взаимодействуя со средой, мхи могут изменять почвенные показатели, такие как температура, влажность, pH (Fenton et al., 2006), оказывать влияние на состав микробных и грибных сообществ (Sedia, Ehrenfeld, 2005). Кроме того, мохообразные чувствительны к изменениям окружающей среды, и их можно рассматривать как индикатор особенностей местообитаний (Ермолаева, Шмакова, 2016; Шафигуллина и др., 2019; Mironov et al., 2020а).
В последние десятилетия в связи с продолжающимся изучением процессов депонирования лесами углерода активно проводятся исследования по оценке прироста мхов и их чистой первичной продукции в зависимости от внешних факторов (Backéus, 1988; Грабовик, 2002, 2003; Гончарова, Беньков, 2005; Гончарова, 2008; Гончарова, Бенькова, 2015; Косых и др., 2017; Grabovik, Nazarova, 2013; Mironov et al., 2020b; Bengtsson et al., 2021). Погодные условия в течение вегетационного периода в значительной мере обуславливают продуктивность мохового покрова (Grabovik, Nazarova, 2013; Костина и др., 2016; Шафигуллина и др., 2019). В частности, влажное и прохладное лето способствует интенсивному росту мхов (Шафигуллина и др., 2019), в то время как относительно высокие температуры и небольшое количество осадков в летний период могут ограничивать их рост (Ермолаева, Шмакова, 2016). В литературе имеются данные, согласно которым линейные приросты Sphagnum fuscum и S. magellanicum, произрастающих в средней тайге Западной Сибири, зависят от суммы годовых и летних осадков, причем годовые осадки имеет большее значение для прироста, чем летние осадки (Косых и др., 2017).
На ростовые процессы мхов большое влияние оказывает температура воздуха и в меньшей степени фотопериод (Mironov et al., 2020b). Так, в Московской области рост мхов начинается весной, когда температура воздуха устанавливается не ниже +5…+6°С. Осенью, при понижении температуры воздуха до этих значений, увеличение размеров останавливается (Костина и др., 2016). В то же время в лесном поясе Хибин побеги мхов видов Hylocomium splendens и Pleurozium schreberi могут продолжать расти и при более низких температурах (Ermolaeva et al., 2013).
Известно, что древесный полог является эдификатором лесных экосистем и влияет на свойства почвы, что может стимулировать развитие мохового покрова. Например, для хвойных лесов Республики Татарстан описано увеличение влажности местообитаний, происходящее в присутствии ели, которое сказалось на максимальном приросте Pleurozium schreberi и, в особенности, на приросте Rhytidiadelphus triquetrus, развивающемся во влажных и освещенных участках еловых редколесий (Шафигуллина и др., 2019).
Методические рекомендации по определению прироста текущего года разных видов мхов встречаются в научных работах разрозненно, и большинство из них предлагают не инвазивные методы того или иного маркирования с измерением прироста через год (Clymo, 1970; Rieley et al., 1979; Zechmeister, 1995).
Цель нашего исследования – оценить годичную продукцию мхов в среднетаежных сосняках кустарничковых Карелии. Для достижения этой цели решались следующие задачи:
- определить годичный прирост встреченных на пробных площадях мхов;
- на основе литературных данных и собственных наблюдений описать комплекс признаков для определения годичного прироста наиболее распространённых в лесных сообществах среднетаёжной зоны Карелии зелёных и сфагновых мхов.
ОБЪЕКТЫ ИССЛЕДОВАНИЯ
Определение массы годичных приростов мхов проводилось в заповеднике «Кивач», расположенном в среднетаежной подзоне Республики Карелия. Для данной территории характерен умеренно континентальный климат с чертами морского. Среднегодовая температура воздуха по многолетним наблюдениям составляет +2.0…+2.5°С, средняя температура января –9…–10°С, июля +16…+17°С. Сумма осадков по многолетним исследованиям составляет 578…618 мм в год (Назарова, 2014, 2015).
Исследования охватывали 15 пробных площадей (ПП), заложенных в сосняках брусничных (СБ) и черничных, произрастающих на подзолах песчаных (СЧ1), и сосняках черничных на подзолах на двучленных отложениях (СЧ2). Сообщества первого типа леса (СБ) характеризовались бедными условиями произрастания, невысоким видовым разнообразием растений и участием кустистых лишайников в напочвенном покрове. Условия местообитания СЧ2 можно описать, как относительно богатые, здесь выше обилие Hylocomium splendens и мхов р. Dicranum (табл. 1).
Таблица 1. Характеристика пробных площадей разных типов леса
| Параметр | СБ | СЧ1 | СЧ2 |
| Тип почвы
|
Подзол иллювиально-железистый
Albic Podzol Arenic |
Подзол иллювиально-железистый
Albic Podzol Arenic |
Подзол иллювиально-гумусовый
Albic Carbic Podzol Abruptic |
| Почвообразующая порода | Флювиогляциальные песчаные отложения
Fluvioglacial sand |
Флювиогляциальные песчаные отложения
Fluvioglacial sand |
Двучленные отложения
Binomial deposit |
| Средний состав древостоя | 10С180 | 10С180–190+ЕедБ | 8С180–1902Е+Б |
| Средний бонитет | III,3 | II,4 | I,3 |
| Относительная
полнота (мин.–макс.) |
0.6–0.9 | 0.8–1 | 1 |
| Количество видов травяно-кустарничкового яруса (мин.–макс.) | 4 | 3–12 | 19–21 |
| Доминанты травяно-кустарничкового яруса (проективное покрытие) | Vaccinium myrtillus (25%)
V. vitis-idaea (15%) |
Vaccinium myrtillus (25%)
V. vitis-idaea (12%) |
Vaccinium myrtillus (35%)
V. vitis-idaea (8%) Calamagrostis arundinacea (6%) |
| Доминанты мохово-лишайникового яруса | Pleurozium schreberi (60%)
Cladonia rangiferina (12%) |
Pleurozium schreberi (70%)
Hylocomium splendens (13%) |
Hylocomium splendens (35%)
Pleurozium schreberi (25%) Dicranum spp. (12%) |
Примечание: СБ – сосняки брусничные, СЧ1 – сосняки черничные на песчаных подзолах, СЧ2 – сосняки черничные на подзолах на двучленных отложениях.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились в полевой сезон 2023 года в первой половине августа. На каждой ПП с помощью рамки 25х25 см закладывались 3–4 учетные площадки. Одна из этих площадок размещалась в межкроновом пространстве, остальные – под кронами сосны. На учетной площадке срезались и разбирались по видам побеги всех мохообразных. Затем, на основании комплекса выявленных в ходе исследования морфологических признаков прироста, на каждом побеге определялся и отделялся годичный прирост. Все образцы годичного прироста с учетной площадки высушивались до абсолютно сухого веса (при 105°С в течение 24 часов) и взвешивались. Таким образом получали данные об абсолютно-сухой массе годичного прироста каждого встреченного вида мха на учетной площадке.
Статистический анализ проводили в программе R (R Core Team, 2023). Всего в статистическом анализе использовали данные с 15 ПП, из них 3 – в сосняке брусничном (СБ), 9 – в сосняке черничном на песчаных подзолах (СЧ1), 3 – в сосняке черничном на подзолах на двучленных отложениях (СЧ2). Влияние кроны сосны было исследовано только для СЧ1. Всего в сообществах данного типа леса было заложено 30 учетных площадок, из них 9 – в межкроновом пространстве, 21 – под кронами сосны. Всего было отобрано и проанализировано 115 образцов мхов (из них 50 – Pleurozium schreberi, 27 – Hylocomium splendens, 38 – Dicranum polysetum). Для сравнения данных между собой применяли критерий Краскела-Уоллиса (аналог дисперсионного анализа для непараметрических данных).
РЕЗУЛЬТАТЫ
К методике определения прироста текущего года у бореальных лесных мхов
Текущий прирост определяется визуально по внешним признакам, общим для многих видов (Корчагин, 1960). В целом определение прироста у всех мхов строится на следующих ключевых признаках: геотропические изгибы стебля, направление прикрепления листьев, цвет стебля.
Геотропические изгибы – это изгибы на стебле (каулидие) мхов, образующиеся в результате восстановления ортотропного направления роста после вдавливания побегов мхов к поверхности субстрата. В природных условиях основным триггером образования геотропических изгибов является давление снежного покрова, такие изгибы называются нивальными (Mironov et al., 2016) (рис. 1, 2).
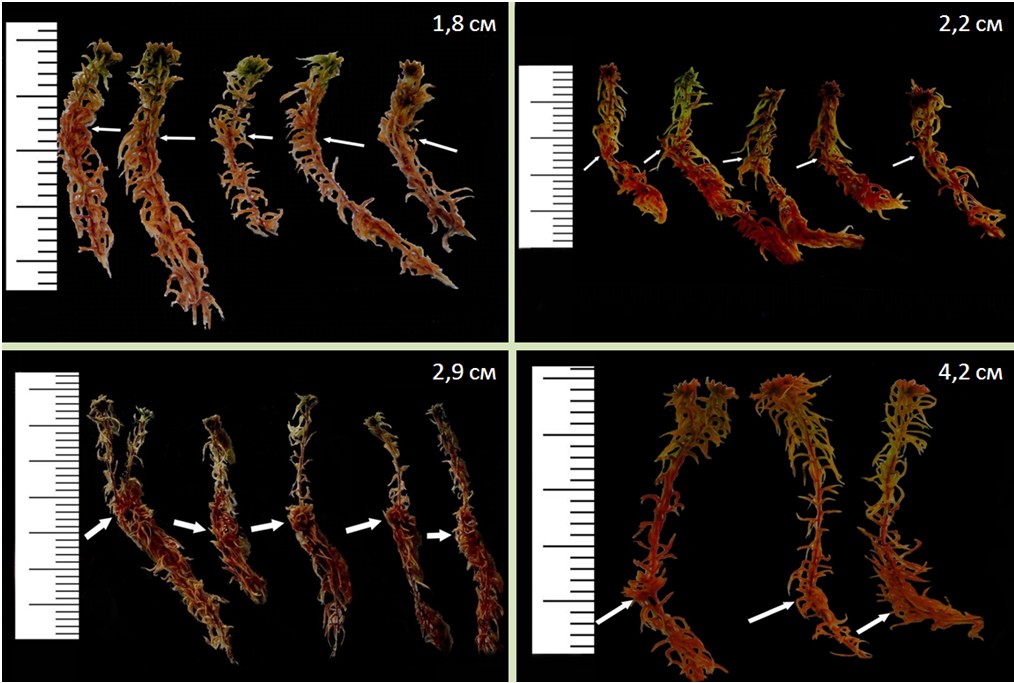
Рисунок 1. Нивальные геотропические изгибы и годичный прирост Sphagnum fuscum
Направление листьев (филлидий). Листья на молодых побегах у большинства видов более или менее прижаты к стеблю, но в течение года, под влиянием внешней среды (капли дождя, снег) угол прикрепления к стеблю меняется (Pouliot et al., 2010). Схематически данное явление отображено на рисунке 2.
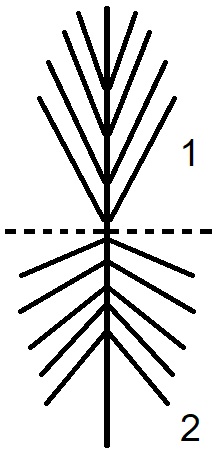
Рисунок 2. Схема прикрепление листьев разного возраста к стеблю у верхоплодных мхов.
1 – прирост текущего года, 2 – прирост прошлого года
Цвет стебля. В большинстве случаев молодые побеги имеют светло-зелёную окраску, в то время как прошлогодние могут быть окрашены в разные оттенки красного и коричневого (Pouliot et al., 2010; Mironov et al., 2016).
Представители родов Polytrichum и Polytrichastrum наиболее просты для определения прироста из всех верхоплодных мхов благодаря хорошо заметному геотропическому изгибу стебля (рис. 3). Также текущий прирост явно виден по цвету, молодой стебель – светло-зелёный, прошлогодний – красно-коричневый.

Рисунок 3. Геотропические изгибы на стебле (каулидии) Polytrichum sp.
Выделение прироста у представителей рода Dicranum также, как правило, не представляет большой трудности благодаря отчётливо видимому изменению угла наклона листьев (рис. 2), особенно хорошо это заметно у Dicranum polysetum (рис. 4).

Рисунок 4. Пример прироста на Dicranum polysetum. Красной линией отмечено начало прироста этого и предыдущего года. Фото сделано в начале лета
Aulacomnium palustre – побеги текущего года зеленого (жёлто-зелёного) цвета и не покрыты плотным войлоком из ризоид (рис. 5)

Рисунок 5. Пример прироста на Aulacomnium palustre. Красной линией отмечено начало прироста. Фото с сайта iNaturalist contributors, iNaturalist (2024). iNaturalist Research-grade Observations. iNaturalist.org. Occurrence dataset https://doi.org/10.15468/ab3s5x accessed via GBIF.org on 2024-06-10. https://www.gbif.org/occurrence/4522719279
У представителей рода Sphagnum наиболее надёжным признаком текущего прироста является геотропический изгиб (Mironov et al., 2016). Если мхи растут в плотной дернине и снежный покров не имеет достаточную массу для вдавливания побегов, геотропические изгибы могут не сформироваться. В этом случае следует пометить растения синтетической ниткой для определения прироста в следующем году (метод перевязок).
Поскольку геотропические изгибы образуются при любом серьезном отклонении побегов, важно учитывать, что сильные дожди, колебания уровня грунтовых вод и другие внешние механические воздействия также могут провоцировать их появление. Такие изгибы могут вводить исследователей в заблуждение, поскольку они не являются маркерами начала вегетационного периода. Чтобы отличать их, важно анализировать прирост в разных частях сфагнового покрова, а также, по возможности, привлекать дополнительные морфологические признаки (Mironov et al., 2016).
Pleurozium schreberi – один из самых сложных мхов для определения прироста. Связано это с тем, что все вышеуказанные признаки выражены слабо, а если растения растут в плотных куртинах, то геотропический изгиб может вовсе отсутствовать. Для определения прироста необходимо изучить комплекс признаков:
1) наличие геотропического изгиба (рис. 1), который у бокоплодных мхов может быть выражен слабо, или не выражен вовсе, но даже если он не выделяется визуально, может быть обнаружен тактильно на стебле;
2) изменение окраски стебля;
3) изменение угла прикрепления листьев к стеблю (рис. 2);
4) изменение формы ветвления (рис. 6, 7).
Если текущий прирост побегов Pleurozium schreberi не возможно определить даже с учётом всех признаков, то необходимо сделать перевязки на стебле, для определения прироста в следующем году (Kostina et al., 2013).
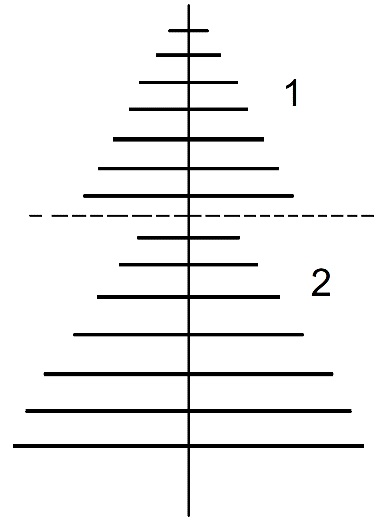
Рисунок 6. Схема изменения формы ветвления на приростах разного года бокоплодного мха. 1 – прирост этого года, 2 – прирост прошлого года (схема примерная, и в реальности различия не так хорошо видны)

Рисунок 7. Пример прироста на Pleurozium schreberi, красной линией отмечено начало прироста этого и предыдущего года. Фото сделано в начале лета
Hylocomium splendens, также известен как «этажный мох». Каждый его новый годичный прирост формирует новую «крону» горизонтальных веточек на стебле (рис. 8). Но нужно учитывать, что в некоторых случаях в благоприятных условиях может формировать два прироста за год.

Рисунок 8. Пример прироста на Hylocomium splendens, красной линией отмечено начало прироста каждого года (на фото их 4 штуки). Фото сделано в начале лета
Прирост Hylocomiadelphus triquetrus (Hedw.) Ochyra & Stebel [Rhytidiadelphus triquetrus] как правило легко определяется по наличию геотропического изгиба, изменению формы ветвления и изменению окраски (рис. 9).

Рисунок 9. Пример прироста на Hylocomiadelphus triquetrus, красной линией отмечено начало прироста этого и предыдущего. Фото сделано в начале лета
Ptilium crista—castrensis и Thuidium sp., как правило, хорошо разделяются на текущий прирост и многолетнюю часть на основании формы ветвления (рис. 10) и геотропического изгиба.
![Рисунок 10. Пример прироста на Ptilium crista-castrensis. Красной линией отмечено начало прироста этого и предыдущего года. Фото с сайта Плантариум. Растения и лишайники России и сопредельных стран: открытый онлайн атлас и определитель растений. [Электронный ресурс] URL: https://www.plantarium.ru/page/view/item/67036.html (дата обращения: 10.06.2024)](https://jfsi.ru/wp-content/uploads/2025/02/Рисунок-10.jpg)
Рисунок 10. Пример прироста на Ptilium crista-castrensis. Красной линией отмечено начало прироста этого и предыдущего года. Фото с сайта Плантариум. Растения и лишайники России и сопредельных стран: открытый онлайн атлас и определитель растений. [Электронный ресурс] URL: https://www.plantarium.ru/page/view/item/67036.html (дата обращения: 10.06.2024)
Для мелких представителей бокоплодных мхов, таких как Sciuro-hypnum sp., Plagiothecium sp., Brachythecium sp., и многих печёночных мхов, если они произрастают рассеянными куртинами на опаде, который каждый год обновляется, вся зелёная часть является текущим приростом.
Довольно часто мохообразные произрастают в смешанных куртинах, которая состоит из видов, приросты которых определяются с разной степенью уверенности. Например, не всегда можно точно определить прирост Pleurozium schreberi, а именно, в плотных куртинах геотропический изгиб и другие признаки могут быть еле заметны, более того сами признаки порой сложно определить (найти на стебле). В таких случаях произрастающие в смеси с ним другие виды, у которых признаки более выражены, например, виды рода Dicranum, могут выступать в качестве маркеров прироста всей куртины, что иногда оказывается хорошей «подсказкой» о среднем приросте мхов в данной области и помогает определить какие признаки выражены на исследуемых трудно определяемых видах, и какова примерная длина годичного прироста.
Запасы фитомассы и годичная продукция
Запасы фитомассы напочвенного покрова, и, в частности, мохово-лишайникового яруса и отдельных видов мхов, в разных типах леса представлены в таблице 2.
Таблица 2. Запасы фитомассы напочвенного покрова по типам леса, г/м2
| СБ | СЧ1 | СЧ2 | |
| мохово-лишайниковый ярус | 295±45 | 239±32 | 110±27 |
| кустистые лишайники | 153±59 | 60±32 | – |
| Pleurozium schreberi | 122±31 | 145±24 | 25±9 |
| Hylocomium splendens | 2±1 | 15±4 | 31±15 |
| Dicranum polysetum | 14±7 | 16±4 | 25±8 |
| надземная фитомасса травяно-кустарничкового яруса | 63±19 | 120±19 | 148±41 |
| подземная фитомасса травяно-кустарничкового яруса | 164±56 | 240±36 | 201±37 |
| общая фитомасса напочвенного покрова | 522±60 | 592±48 | 458±46 |
Примечание: указано среднее значение и стандартная ошибка среднего.
Годичная продукция преобладающих в напочвенном покрове зеленых мхов в зависимости от типа леса представлена на рисунке 11. Для Pleurozium schreberi годичная продукция достоверно выше в сосняках брусничных и черничных на песчаных подзолах по сравнению с сосняком черничным на двучленных отложениях. Для Hylocomium splendens и Dicranum polysetum годичная продукция, напротив, минимальна в сосняке брусничном, но эти значения значимо не различаются в сравнении с сосняками черничными (рис. 11).
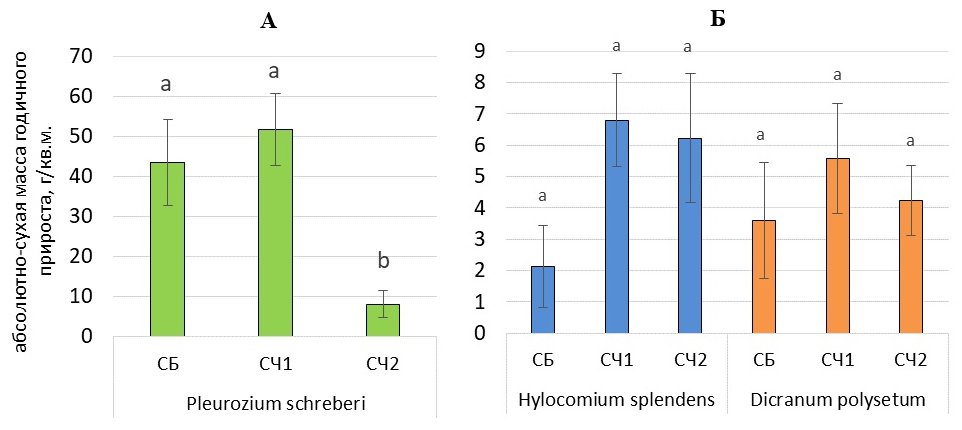
Рисунок 11. Годичная продукция разных видов зеленых мхов в зависимости от типа леса, г/м2: А – Pleurozium schreberi, Б – Hylocomium splendens, Dicranum polysetum (показаны средние значения и стандартная ошибка среднего). Обозначения: СБ – сосняк брусничный, СЧ1 – сосняк черничный на песчаных подзолах, СЧ2 – сосняк черничный на двучленных отложениях. Латинскими буквами обозначены достоверные различия между разными типам леса
Масса годичного прироста зеленых мхов также различалась в разных зонах фитогенного поля. Для Pleurozium schreberi годичная продукция под кроной сосны была в два раза выше по сравнению с межкроновым пространством (табл. 3). Однако даже эти различия статистически не значимы при данной выборке.
Таблица 3. Годичная продукция зеленых мхов в разных зонах фитогенного поля, г/м2
| Вид зеленого мха | Межкроновый участок | Подкроновый участок |
| Pleurozium schreberi | 29.9±9.3 | 61.2±11.8 |
| Hylocomium splendens | 7.7±1.9 | 5.8±2.4 |
| Dicranum polysetum | 3.6±1.1 | 4.2±1.1 |
Отмечено большое варьирование значений годичного прироста зеленых мхов, что не позволило выявить достоверные различия между выборками.
В среднем доля годичной продукции от общей массы мха составляет 0.38±0.02 для Pleurozium schreberi, 0.26±0.02 – для Hylocomium splendens и 0.25±0.02 – для Dicranum polysetum.
ОБСУЖДЕНИЕ
В результате исследования выявлено, что в обследованных среднетаежных сосновых лесах основной вклад в суммарную годичную продукцию мохового покрова вносит Pleurozium schreberi, в среднем 41.4±6.3 г/м2, что в целом соответствует литературным данным для лесов таежной зоны (Шпак, Шмакова, 2010; Осипов, 2017). Наибольшая годичная продукция для данного вида отмечена в сосняках на песчаных подзолах, наименьшая – в высокополнотных и высокобонитетных сосняках черничных с участием ели и березы в древостое и относительно большим участием злаков в напочвенном покрове.
Несмотря на большую разницу в значениях годичной продукции Pleurozium schreberi в разных зонах фитогенного поля, это различие статистически не значимо. Также не удалось выявить различия в годичном приросте по массе между Hylocomium splendens и Dicranum polysetum. Высокое варьирование значений вызвано, вероятно, большой мозаичностью мохово-лишайникового яруса в сосновых лесах. Для лесных сообществ лесостепной зоны указывается, что линейные приросты мха Hylocomium splendens под кронами деревьев и в межкроновом пространстве достоверно не различаются. В то же время прирост по массе под кронами деревьев значимо выше, чем в межкроновом пространстве (Гончарова, Бенькова, 2015). Противоположные результаты были получены при изучении годичной продукции мхов в лесном поясе Хибин: наибольшие значения годичного прироста Hylocomium splendens и Pleurozium schreberi были характерны для просветов крон деревьев, что авторы связывают с более благоприятными условиями для роста (Ermolaeva et al., 2013; Ермолаева, Шмакова, 2016).
ЗАКЛЮЧЕНИЕ
На основе литературных данных и собственных наблюдений выявлены ключевые морфологические признаки годичного прироста у основных представителей зелёных и сфагновых мхов, произрастающих в сосняках брусничных и черничных среднетаёжной зоны Карелии. Определить годичный прирост мхов можно по геотропическому изгибу стебля, изменению направления роста листьев и изменению цвета стебля по сравнению с многолетними частями растения.
В результате проведённого исследования выявлено, что основной вклад в суммарную годичную продукцию мохового покрова обследованных сосновых лесов вносит Pleurozium schreberi (41.4±6.3 г/м2). Средние значения прироста текущего года для Hylocomium splendens и Dicranum polysetum составляют 5.9±1.1 и 4.8±1.1 г/м2 соответственно. В сосняках черничных на песчаных почвах годичная продукция Pleurozium schreberi под кроной сосны в два раза выше по сравнению с межкроновым пространством (61.2±11.8 и 29.9±9.3 г/м2 соответственно). Однако стоит учитывать, что данные результаты измерения годичной продукции получены всего за один год наблюдений. Для получения более точных значений годичной продукции для разных видов мхов и оценки её варьирования необходимы измерения в течение нескольких лет.
ФИНАНСИРОВАНИЕ
Работа выполнена в рамках реализации важнейшего инновационного проекта государственного значения «Разработка системы наземного и дистанционного мониторинга пулов углерода и потоков парниковых газов на территории Российской Федерации, обеспечении создания системы учета данных о потоках климатически активных веществ и бюджете углерода в лесах и других наземных экологических систем» (рег. № 123030300031-6).
СПИСОК ЛИТЕРАТУРЫ
Гончарова И. А. Изменчивость годичного прироста и чистой продукции зелёного мха в тундровых редколесьях западного Таймыра // Лесоведение. 2008. № 3. С. 76–78.
Гончарова И. А., Беньков А. В. Динамика приростов зелёных мхов в лесоболотных комплексах юга Западной Сибири // Лесоведение. 2005. № 1. С. 43–51.
Гончарова И. А., Бенькова А. В. Влияние погодных факторов на динамику годичного прироста и чистой продукции Hylocomium splendens в лесных сообществах лесостепной зоны Хакасии // Сибирский лесной журнал. 2015. № 6. С. 54–61.
Грабовик С. И. Динамика годичного прироста у некоторых видов Sphagnum L. в различных комплексах болот Южной Карелии // Растительные ресурсы. 2002. Т. 38. Вып. 4. С. 62–68.
Грабовик С. И. Динамика продуктивности ценопопуляций сфагновых мхов южной Карелии // Ботанический журнал. 2003. Т. 88. № 4. С. 41–48.
Ермолаева О. В., Шмакова Н. Ю. Рост и накопление массы Hylocomium splendens в лесном поясе Хибин // Ученые записки Петрозаводского государственного университета. 2016. № 8 (161). С. 40–45.
Корчагин А. А. Определение возраста и длительности жизни мхов и печеночников // Полевая геоботаника. 1960. Т. 2. С. 279–314.
Костина М. В., Сафронова Г. А., Барабанщикова Н. С. Влияние погодных и микроклиматических условий на величину годичных линейных приростов и строение побеговых комплексов некоторых бокоплодных мхов Московской области // Бюллетень Московского общества испытателей природы. Отдел биологии. 2016. Т. 121. Вып. 1. С. 45–53.
Косых Н. П., Коронатова Н. Г., Лапшина Е. Д., Филиппова Н. В., Вишнякова Е. К., Степанова В. А. Линейный прирост и продукция сфагновых мхов в средней тайге Западной Сибири // Динамика окружающей среды и глобальные изменения климата. 2017. Т. 8. № 1 (15). С. 3–13.
Назарова Л. Е. Атмосферные осадки в Карелии // Труды КарНЦ РАН. 2015. № 9. С. 114–120.
Назарова Л. Е. Изменчивость средних многолетних значений температуры воздуха в Карелии // Известия Русского географического общества. 2014. Т. 146. № 4. С. 27–33.
Осипов А. Ф. Запасы и потоки органического углерода в экосистеме спелого сосняка черничного средней тайги // Сибирский лесной журнал. 2017. № 2. С. 70–80. DOI: 10.15372/SJFS20170208.
Телеснина В. М., Подвезенная М. А., Сорокин А. С., Мешалкина Ю. Л. Оценка биомассы хвойно-лиственных лесов на примере УОПЭЦ МГУ «Чашниково» // Вестник Московского университета. Серия 17. Почвоведение. 2024. Т. 79. № 2. С. 37–45. DOI: 10.55959/MSU0137-0944-17-2024-79-2-37-45.
Шафигуллина Н. Р., Каржавкина Е. Н., Зиятдинова З. Ф. Влияние различных факторов на продуктивность мохового покрова хвойных лесов Республики Татарстан // Экосистемы. 2019. № 17. С. 44–55.
Шпак О. В., Шмакова Н. Ю. Первичная продукция мхов в Хибинах (Кольский полуостров) // Растительные ресурсы. 2010. Т. 46. № 2. С. 42–50.
Backéus I. Weather variables as predictors of Sphagnum growth on a bog // Ecography. 1988. Vol. 11. P. 146–150.
Bengtsson F., Rydin H., Baltzer J. L., Bragazza L., Bu Z. J., Caporn S. J., Granath G. Environmental drivers of Sphagnum growth in peatlands across the Holarctic region // Journal of Ecology. 2021. № 109. P. 417–431.
Clymo R. S. The growth of Sphagnum: methods of measurement // The Journal of Ecology. 1970. P. 13–49.
Cornelissen J. H. C., Lang S. I., Soudzilovskaia N. A., During H. J. Comparative cryptogam ecology: a review of bryophyte and lichen traits that drive biogeochemistry // Annals of botany. 2007. Vol. 99. P. 987–1001. DOI: 10.1093/aob/mcm030
Ermolaeva O. V., Shmakova N. Yu., Lukyanova L. M. On the growth of Polytrichum, Pleurozium, and Hylocomium in the forest belt of the Khibiny Mountains // Journal of Bryology. 2013. Vol. 22. P. 7–14.
Fenton N. J., Légaré S., Bergeron Y., Paré D. Soil oxygen within boreal forests across an age gradient // Canadian Journal of Soil Science. 2006. Vol. 56. P. 1–9.
Grabovik S. V., Nazarova L. Linear increment of Sphagnum mosses on Karelian mires (Russia) // Arctoa. 2013. Vol. 22. P. 15–22.
iNaturalist contributors, iNaturalist. 2024. iNaturalist Research-grade Observations. iNaturalist.org. Occurrence dataset https://doi.org/10.15468/ab3s5x accessed via GBIF.org on 2024-06-10. URL: https://www.gbif.org/occurrence/4522719279 (дата обращения 06.10.2024).
Kostina M. V., Safronova G. A., Agapov P. A. On the growth of Pleurozium schreberi (Bryophyta) in Moscow Province //Arctoa. 2013. Vol. 22. P. 15–22.
Mironov V. L., Grabovik S. I., Ignashov P. A., Kantserova L. V. Geotropic curvatures of Sphagnum: environmental features of their genesis and trial application for estimation shoot length increment // Arctoa. 2016. Vol. 25. P. 353–363. DOI: 10.15298/arctoa.25.27.
Mironov V. L., Kondratev A. Y., Mironova A. V. Growth of Sphagnum is strongly rhythmic: contribution of the seasonal, circalunar, and third components // Physiologia Plantarum. 2020a. Vol. 168. № 4. P. 765–776.
Mironov V. L., Kondratev A. Y., Mironova A. V. Sphagnum growth as an indicator of wavelength-specific UV-B penetration through the ozone layer // Ecological Indicators. 2020b. Vol. 116. P. 106430.
Pouliot R., Marchand-Roy M., Rochefort L., Gauthier G. Estimating moss growth in arctic conditions: a comparison of three methods // The Bryologist. 2010. Vol. 113. No 2. P. 322–332.
R Core Team. 2023. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. URL: https://www.R-project.org/ (дата обращения 06.101. 2024).
Rieley J. O., Richards P. W., Bebbington A. D. L. The ecological role of bryophytes in a North Wales woodland // The Journal of Ecology. 1979. P. 497–527.
Sedia E. G., Ehrenfeld J. G. Differential effects of lichens, mosses, and grasses on respiration and nitrogen mineralization in soils of the New Jersey Pinelands // Oecologia. 2005. Vol. 144. № 1. P. 137–147. DOI: 10.1007/s00442-005-0037-0.
Zechmeister H. G. Growth rates of five pleurocarpous moss species under various climatic conditions // Journal of bryology. 1995. Vol. 18. №. 3. P. 455–468.
Рецензент: д.б.н. Шмакова Н. Ю.





