- DOI: 10.31509/2658-607x-202583-173
- УДК 57. 574.4
Органопедогенез – незамеченный базовый процесс почвообразования
![]()
© 2025 О. Г. Чертов*, Н. В. Лукина
Центр по проблемам экологии и продуктивности лесов им. А. С. Исаева Российской
академии наук, Россия, 117997 Москва, ул. Профсоюзная, 84/32, стр. 14
*E-mail: ochertov@rambler.ru
Поступила в редакцию: 21.05.2025
После рецензирования: 01.07.2025
Принята к печати: 15.07.2025
Предлагается концепция органопедогененеза – более века незамечаемого активного процесса общего педогенеза, который отражает весомую роль биологического фактора почвообразования в русле представлений В. И. Вернадского о ведущем значении организмов во всех природных процессах в континентальной части биосферы. Представлена методология отражения роли органопедогенеза в существующих почвенных классификациях путем включения в их систематику типов аккумуляции и трансформации органического вещества почв совместно с типами органопрофилей. Включение этих компонентов придаст классификации почв динамичность и увеличит практическую значимость почвоведения, которые особенно важны для принятия управленческих решений в настоящее время в условиях быстрых негативных изменений природной среды.
Ключевые слова: биологический фактор почвообразования, органопрофиль, аккумуляция и трансформация ПОВ, формы гумуса
Идея о масштабной и активной роли биологического фактора в формировании и развитии почв уже много лет близка последователям учения о биосфере В. И. Вернадского (Родин, Базилевич, 1965; Будыко, 1984; Ковда, 1985; Одум, 1986; Глазовская, 1988; Горшков, 1995). Однако до настоящего времени эта роль не проанализирована на основе новых данных и не обобщена с точки зрения вклада в педогенез (Эволюция почв …, 2015; Эволюция, функционирование…, 2020). Именно обсуждению значимости биологического фактора в генезисе и особенно классификации почв и посвящен представленный ниже материал[1]. Концепция биосферы В. И. Вернадского (1926, 1994) служит теоретическим фундаментом научно обоснованного природопользования, что хорошо известно и принимается научным сообществом. Она появилась вслед становлению почвоведения как самостоятельной науки о Земле. Биосфера – это тонкая оболочка планеты с активным «живым веществом» (биомассой сообществ организмов), являющимся мощным фактором эволюции земной коры на геологических временах и активным реактором, в котором в планетарном масштабе протекают современные биогеохимические процессы в приземном слое атмосферы, на поверхности литосферы и в гидросфере. При этом биосфера имеет свои механизмы обратной связи с этими тремя сферами и пути восстановления собственной структуры и функций после нарушений. Педосфера – это особый активный пограничный слой в структуре биосферы между живыми организмами и континентальной частью литосферы, то есть это почвенный покров планеты Земля (Карпачевский, 1981; Карпачевский и др., 1984; Вернадский, 1994; Добровольский и др., 2010). Это тонкая плёнка воздействия живых организмов и продуктов их жизнедеятельности на верхнюю зону выветривания горных пород земной коры, обеспечивающая эффективное функционирование организмов биосферы за счёт биотического круговорота вещества и биогенных потоков энергии. Она обладает буферностью, которая поддерживает стабильность биологических сообществ и самой почвы. Наличие в педосфере активной биоты и продуктов её жизнедеятельности чётко отграничивает почву от литосферы (Шоба, 1988; Добровольский, 2009; Бахмет, 2015; Таргульян, 2019).
Современное почвоведение базируется на представлениях В. В. Докучаева (Докучаев, 1883; Сибирцев, 1899; Ярилов, 1904) о пяти факторах почвообразования (климат, рельеф, почвообразующие породы, организмы, время) к которым теперь ещё добавлен антропогенный фактор. При этом изначально постулировалась равнозначность действия этих факторов. Эта базовая концепция до настоящего времени признана основой генетического почвоведения. Однако уже П. А. Костычев (1886) подчеркивал ведущую роль растительности в формировании почв, что не укладывалось в принцип равнозначности влияния факторов почвообразования. Надо полагать, что справедливость этих постулатов связана с оперируемыми временными периодами: в масштабе тысячелетий справедливо классическое определение, тогда как на интервалах нескольких десятилетий и столетий, в течение которых не меняются почвообразующие породы и рельеф, вполне обоснованной является точка зрения П. А. Костычева о вкладе растительности как представителя самого динамичного, биотического фактора почвообразования хотя мощность его влияния безусловно зависит и от других составляющих педогенеза. Поскольку в настоящее время наша планета испытывает быстрые изменения климата, этот фактор также относится к динамичным, и влияние современных климатических изменений на почвы, наиболее ярко проявляющихся с 80-х годов прошлого столетия, также проявляется через биологический фактор, прежде всего, растительность и почвенную биоту (Лукина, 2025).
Рассмотрение почвообразования в сочетании не только с первичной экогенетической сукцессией растительности [по В. Н. Сукачёву, это «эндоэкогенез» (Сукачев, Дылис, 1964)], но и с разнообразными вторичными сукцессиями лесной растительности, вызванными различными нарушениями (рубки, пожары, шторма с образованием ветровально-почвенных комплексов, промышленное загрязнение и сельскохозяйственное использование), позволяет заключить, что и при первичной сукцессии, и в ходе вторичных сукцессий выражен тренд накопления почвенного органического вещества (ПОВ) (Дюшофур, 1979; Чертов, 1981, 1990; Карпачевский и др., 1984; Одум, 1986; Разумовский, 1999; Лукина и др., 2010, 2019; Данилов и др., 2024; Emmer, 1995; Nadporozhskaya et al., 2006; Kalinina et al., 2009). В оставшихся на небольших площадях старовозрастных малонарушенных лесах (Лукина и др., 2023), сформировавшихся в ходе непрерывного оборота поколений ключевых видов-эдификаторов (Смирнова, 1998; Смирнова, Торопова, 2008, 2016), в которых длительное время (сотни лет) преобладали естественные процессы и образовался абсолютно-разновозрастный древостой (Дыренков, 1984), можно ожидать самого высокого уровня накопления ПОВ. Причем, процесс накопления ПОВ представлен нелинейным трендом с выраженными пиками и падениями, обусловленными соотношениями процессов аккумуляции и потерь органического вещества за счёт эмиссии углекислого газа из почв и выноса органического вещества с почвенными водами в ходе естественных нарушений.
Более того, общий динамический тренд естественного почвообразования представляет собою последовательную эутрофикацию почвы с концентрированием не только углерода и азота органического вещества, но и других элементов-биофилов (Чертов, Разумовский, 1980; Чертов, 1990), что на временах столетий сукцессионных смен растительности без нарушений может прослеживаться даже в подзолах и торфяных почвах (Чертов, 1981, 1990). Из этого следует, что «теория единого почвообразовательного (дернового) процесса» В. Р. Вильямса (цит. по Вильямс, 2025) всё же отражает общий тренд эутрофикации почв в ходе экогенетических сукцессий.
Следует признать, что если мы принимаем доминирование биологического фактора основополагающим в формировании почв в русле биосферных представлений В. И. Вернадского (Пономарева, 1964; Родин, Базилевич, 1965, Чертов, Разумовский, 1980, 1981; Карпачевский и др., 1984; Лукина и др., 2010, 2022, Lukina et al., 2019), то аккумуляция ПОВ служит базовым системообразующим процессом, отличающим почвы от геологического образования в наземных и мелководных ландшафтах разного уровня (биомах, экосистемах/биогеоценозах, экотопах и др.). Присутствие даже в самом незначительном количестве почвенного (а не вообще любого) органического вещества – это признак почвы в разных стадиях своего развития. Отсутствие ПОВ – это признак геологического или антропогенно-модифицированного субстрата или горной породы, но не почвы. Наличие ПОВ при отсутствии минерального материала свидетельствует о том, что это почва, но на органогенных почвообразующих породах, которые изначально формировались как на минеральной матрице, так и в водной среде. С другой стороны, даже на массивных горных породах может формироваться почва-плёнка с крипто-микозным ПОВ и нано-процессами его аккумуляции, приводящими затем к появлению лишайников (Chertov et al., 2004; Lessovaia et al., 2008) и развитию процессов почвообразования в ходе первичной экогенетической сукцессии.
Следует подчеркнуть, что основным твердофазным морфологически чётко определимым макро-продуктом влияния биоты на почвообразование является ПОВ, которое изучают много и обстоятельно (Трофимов и др., 1986; Орлов и др., 1996; Чуков, 1997; Семенов, Когут, 2015; Piccolo et al., 2019; и многие другие). Но, к сожалению, исследования строения, структуры и функции ПОВ в начале ХХI в. в теоретическом отношении возвращаются на позиции столетней давности практически с отказом от научного багажа середины ХХ в. (Семенов, Когут, 2015; Lehmann, Kleber, 2015). Фракционирование ПОВ фактически редуцировано до двух физических фракций: твердых органических частиц (particulate organic matter, РОМ) и минерально-ассоциированного органического вещества (mineral-associated organic matter, МАОМ) (Семенов и др., 2023), а игнорирование прошлого научного багажа доходит до отказа от термина «гумус» (Lehmann, Kleber, 2015). Последнее является крайностью, которую в своё время З. А. Ваксман (1937) себе не позволил: темноокрашенное органическое вещество в верхних горизонтах почв с историческим термином «гумус» является объективной реальностью, как бы ни менялись наши представления о его химическом составе и физической структуре. С одной стороны, налицо практически единодушное признание выдающейся роли «живого вещества биосферы» в формировании зелеными растениями и материалом их переработки почвенной биотой ПОВ, как масштабного продукта в процессе почвообразования (Пономарева, 1964; Карпачевский и др., 1981; Вернадский, 1994; Добровольский и др., 2010; Семенов, Когут, 2015; Чертов, 2025; и др.). С другой стороны, на классификационном уровне отмечается аморфная позиция в отношении конкретизации роли ПОВ в педогенезе: в генетических классификациях почв не наблюдается тенденции к отражению разнообразия морфологических форм ПОВ. Даже после декларирования В. Р. Вильямсом (цит. по Вильямс, 2025) в 30-х годах ХХ в. «диалектически обоснованной» концепции биологической эволюции почвы так и не произошло какого-либо существенного сдвига в советской почвенной классификации (за исключением введения в классификацию подтипа «дерново-подзолистых почв»).
Существующие классификации почв
В существующих почвенных классификациях незаменимая роль ПОВ в генезисе почв представлена очень слабо. Так, в Международной системе почвенной классификации WRB (Мировая …, 2024; IUSS Working Group WRB, 2022), вырисовывается следующая картина:
1) Почвы называются по доминирующему признаку (часто просто по цвету) с обязательным использованием только устойчивых параметров минеральных горизонтов, поскольку «свойства верхнего горизонта могут быстро изменяться во времени, поэтому в системе WRB они используются для диагностики только в редких случаях» (Мировая …, 2024, стр. 25). То есть свойства почвы, отражающие её как динамическую развивающуюся систему, считаются мешающими классифицированию.
2) В названиях «реферативных почвенных групп» обращает на себя внимание «абиологичность» классификации по отражению свойств ПОВ, как продукта трансформации растительных остатков всей почвенной биотой. Так, из 32 наименований «реферативных почвенных групп» («отделы» в российской классификации) только четыре соотнесены с ПОВ (чернозёмы, Phaeozems, Umbrisols и органические почвы). Более того, из 294 «квалификаторов» (генетических признаков в форме уточняющих прилагательных к наименованию почвы) только 14 соотнесены с ПОВ. Это было бы правильно, если бы речь шла не о почве, а о классификации форм выветривания четвертичных отложений и горных пород.
В отличие от европейского подхода, почвенная классификация России (Классификация и диагностика …, 2004) является более биологической и экологической: в ней из 52 диагностических горизонтов 24 (46%!) базируется на признаках ПОВ (Полевой определитель …, 2008). Однако, в целом в этих почвенных классификациях не прослеживается стремления к обязательному таксономическому отражению во всех таксонах двух базовых твердых фаз почвенной системы: почвенного органического вещества, как продукта почвообразования, и минеральной основы всей почвы. В. И. Вернадский (1994) определял почву как «биокосное тело», т.е. двухкомпонентное, состоящее из активной биологической и инертной косной (в мягком значении этого слова) части, которая относится к минеральной матрице. Однако, до настоящего времени в почвенных классификациях доминирует «косная» составляющая.
Несмотря на то, что идея классификационного отражения роли гумуса лежит буквально на поверхности более ста лет, ПОВ оказалось пасынком существующих генетических классификаций со скромным местом в общей теории педогенеза (Соколов, 1997; Таргульян, 2019).
Органопедогенез
В связи с вышеизложенными теоретическими положениями предлагается ввести понятие «органопедогенез»[2], как важнейший системообразующий процесс формирования и эволюции почвы, который путём аккумуляции и трансформации ПОВ автотрофными и гетеротрофными организмами влияет на все функции и общий генезис почвенной системы, безусловно, с учётом и под влиянием действия всех других классических факторов почвообразования. Это справедливо как в отношении традиционно понимаемого генезиса с эволюцией всего почвенного профиля, так и при переходе к оперированию непрерывными процессами: геохимическими, гидрологическими и, более всего, собственно биологическими, которые определяют обратную связь от почвы ко всей биоте в биосфере, с учётом всего биоразнообразия, продуктивности естественных и антропогенных фито-, зоо- и микробоценозов и роли почвы в глобальном круговороте углерода. То есть органопедогенез – это многокомпонентный базовый почвенный процесс формирования органогенных и гумусово-аккумулятивных горизонтов и всего органопрофиля почвы. Однако, до настоящего времени органопедогенез – это фактически отсутствующий аспект в учении о генезисе почв, который является упущенной концепцией в почвенных генетических классификациях. Семантически идея органопедогенеза уже присутствует в генетическом почвоведении, например, в термине органо-«генные» почвы. Каковы же перспективы реализации и развития концепции органопедогенеза?
Компоненты органопедогенеза
На основе изложенных выше положений представляется возможным выделить два морфологически выраженных результата органопедогенеза: а) различающиеся типы аккумуляции и трансформации ПОВ, б) различающиеся типы органопрофилей всей почвенной системы. При этом обнаруживается, что для развития этих аспектов имеется достаточно хорошо проработанный научный базис. Во-первых, в лесном почвоведении существует подход, который полностью вкладывается в концепцию органопедогенеза. Это классификация и диагностика «форм/типов гумуса» на основе морфологически выраженных признаков аккумуляции и трансформации ПОВ в органическом и гумусово-аккумулятивном горизонтах, которая появилась сначала на западе (Müller, 1887; Kubiena, 1953; Wilde, 1958; Duchaufour, 1965) и далее последовательно развивалась в Германии (Broll et al., 2006; Wachendorf et al., 2023), США и Канаде (Wilde, 1958; Green et al., 1993; Klinka, 1997), в последнее время интенсивно изучалась в Европейском Союзе (Ponge, 2003; Humusica …, 2018; Kõlli, Köster, 2018; Zanella еt аl., 2018, 2022) и продуктивно развивалась в России (Крюденер, 1916; Тюрин, Пономарева, 1940; Благовидов, Бурков, 1959; Чертов, 1974, 1981, 2025; Чертов, Надпорожская, 2018; Chertov, Nadporozhskaya, 2017). Во-вторых, это концепция почвенных органопрофилей (Гришина, 1986; Шоба, 1988; Миньковский, Шоба, 1994, 1995; Бахмет, 2015).
Типы аккумуляции и трансформации ПОВ (формы гумуса)
Систематика и диагностика форм гумуса «покоится на трех китах»: «грубый гумус» («мор» на западе) с развитой подстилкой, «мягкий гумус» («мулль») с развитым гумусовым горизонтом при маломощной подстилке и переходный тип «модер», у которого в наличии оба горизонта. Термин «мор» (mor), грубый гумус, в своё время критиковался в США Уайльдом (Сергей Александрович Вильде) (Wilde, 1958) в связи с его значением в русском языке. Эта триада также прослеживается в классификации форм гумуса органогенных почв по градиенту увлажнения вплоть до трех типов торфяников, а также имеет разновидности в почвах луговых и степных экосистем (Humusica …, 2018). В российской классификации форм гумуса имеется большое количество переходных типов (рис. 1). Ещё по предложению И. В. Тюрина и В. В. Пономаревой (1940) с последующим широким использованием на Северо-Западе РФ (Благовидов, Бурков, 1959; Чертов, 1966, 1981; Чертов и др., 1974, 1978) названия форм/типов гумуса обозначены прилагательными к слову «почва». Все эти формы/типы гумуса различаются по морфологическим и физико-химическим параметрам, которые определяются прежде всего качеством поступающего в почву опада: наземный и корневой опад хвойных пород с низким содержанием азота и других элементов питания приводит к формированию медленно разлагающегося грубого гумуса, тогда как опад лиственных деревьев с более высоким качеством опада, то есть с повышенным содержанием этих элементов питания (Лукина и др., 2008, 2019, 2021; Биоразнообразие …, 2021), позволяет почвенной фауне и микроорганизмам быстро его перерабатывать с образованием муллевой формы гумуса. Поэтому формы гумуса чётко связаны с характером и интенсивностью гумификации, а также с составом, продуктивностью и биоразнообразием лесной растительности (Чертов, 1981; Федорчук и др., 2005; Чертов, Надпорожская, 2018). В свою очередь, разнообразный состав растительности с разным качеством опада, то есть с разным содержанием различных соединений азота, фосфора, кальция, микроэлементов является стартёром формирования различных форм гумуса.
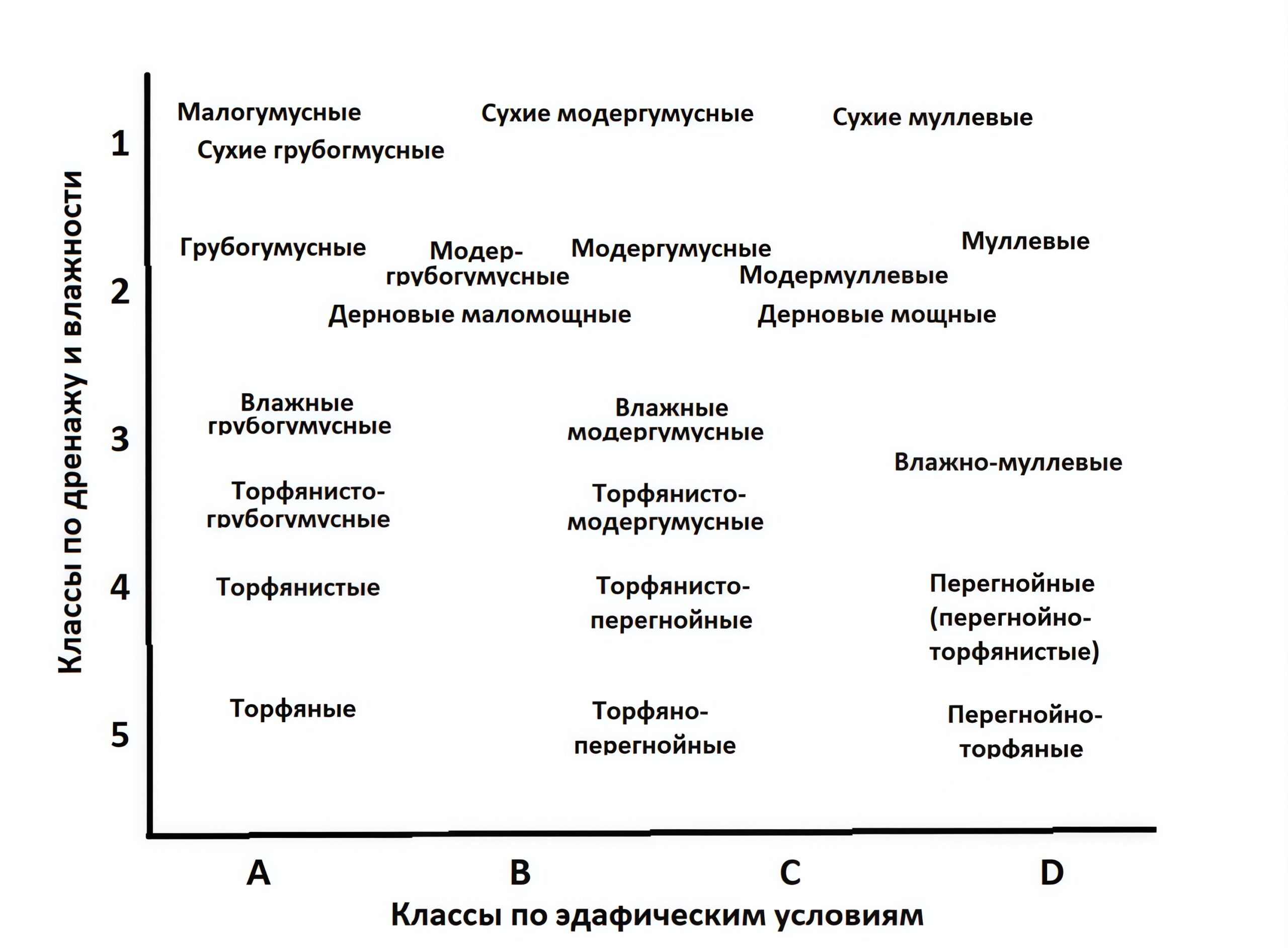
Рисунок 1. Ординация типов аккумуляции ПОВ (форм гумуса) лесных почв по классам богатства (А, бедные – D, богатые) и влажности (1, сухие – 5, постоянно избыточно переувлажненные) типов лесорастительных условий лесной зоны Европейской России (Чертов, 1981; Чертов, Надпорожская, 2018; Chertov, Nadporozhskaya, 2017). Классы эдафических условий — принятые в лесоведении градации богатства (плодородия) почв.
Эта классификация развивалась в рамках лесного почвоведения, но так и не была воспринята в генетическом почвоведении. Причиной этого могут быть следующие обстоятельства: а) концепция форм гумуса развивалась в период интенсивного распространения идей В. В. Докучаева о почвообразовании и была оттеснена в разряд прикладного, лесного почвоведения; б) произошла ошибка в переводе немецкого термина Humus Form (для горизонтов О и Аh) как формы лесных подстилок (нем. Humusdecke), а не форм гумуса. Это привело к развитию в российском почвоведении типологии подстилок (Шумаков, 1958; Карпачевский, 1981; Богатырев и др., 2004), и содержательное изучение лесных подстилок никогда не снималось с повестки дня в лесной экологии и почвоведении. Морфологическая классификация форм гумуса эволюционировала более века параллельно мейнстриму генетического почвоведения.
Теоретической основой выделения типов аккумуляции и трансформации ПОВ как форм гумуса является ныне забытая «эдафология» (Чертов и др., 2018), с которой начиналось формирование почвоведения: в историческом аспекте она вторая после «агрогеологии» до появления принципов генетического почвоведения В. В. Докучаева. В эдафологии в понимании российской школы почва рассматривается как подземная среда обитания авто- и гетеротрофных организмов, прежде всего растений. В настоящее время этот подход ближе всего к экологическому почвоведению, лесоведению и ландшафтоведению. Отметим, что все существующие математические модели динамики ПОВ являются эдафологическими (Komarov et al., 2017).
В России имеется успешный опыт включения форм гумуса в генетическую классификацию почв при картировании лесных почв и в лесотипологических исследованиях (Чертов и др., 1978; Чертов, 1981; Дыренков, 1984; Федорчук и др., 2005). В общей сложности формы гумуса были включены в систематический список почв на картах масштаба от 1:10000 до 1:100000 семи лесхозов площадью 350 тыс. га (Чертов, 1981).
На западе интерес к классификации форм гумуса был особенно интенсивен в начале ХХI в. в связи с систематизацией имеющихся данных и даже с заменой традиционного термина «форма гумуса» на humipedon и призывом к развитию «биологического почвоведения» (Humusica …, 2018), поскольку попытки интеграции в генетическую классификацию WRB не увенчались успехом, за исключением Франции, в которой формы гумуса были включены в почвенную таксономию (Дюшофур, 1979). В западных странах классификации и полевые определители форм гумуса включены в руководства по картографированию почв и лесоустроительные инструкции (Малышева и др., 2022), что так же было сделано и в России (Чертов, 2025).
Почвенные органопрофили
Формы гумуса представляют собою классификацию только типов аккумуляции и трансформации ПОВ в зоне активного биотического круговорота, но не всего почвенного профиля (Чертов, Надпорожская, 2018; Zanella et al., 2022). Почвенный органопрофиль по определению их авторов – это другое понятие, относящееся к «закономерному сочетанию и распределению ПОВ по всему генетическому профилю почвы с характеристикой его специфических особенностей» (Гришина, Орлов, 1978; Гришина, 1986). Это понятие было ориентировано на более всестороннюю диагностику почв с позиций генетического почвоведения. С экологических позиций концепция органопрофилей использовалась О. Н. Бахмет (2015), которая трансформировала её для расширения типов аккумуляции ПОВ, то есть форм гумуса. Однако, это не вполне соответствует определению Л. А. Гришиной, поскольку одинаковые типы аккумуляции ПОВ (формы гумуса) могут формироваться в почвах с разным гумусовым профилем. Например, грубогумусная форма может быть и в почвах с монотонным убыванием ПОВ по профилю, и в почвах с гумусово-иллювиальным (альфегумусовым) и даже с маломощным недифференцированным профилем на скальном основании. Это так называемые «почвы-аналоги» по Ф. Дюшофуру (1979).
Полный органопрофиль почвы отражает выход ПОВ из зоны аккумуляции и биотического круговорота (формы гумуса) в зону биогеохимических циклов с транзитом, выносом, элювиированием и с синхронной трансформацией минеральной матрицы (Перельман, 1975; Фокин, 1975; Глазовская, 1988), вызванной влиянием биологического фактора. То есть, органопрофиль по Л. А. Гришиной (1986) – это прежде всего генетическое, а не эдафо-экологическое понятие. В теории морфогенеза почв С. А. Шобы (1988) типология почвенных профилей не была разработана: у автора это в основном систематизация микроморфологических признаков процессов трансформации минеральной матрицы.
Ниже предлагается авторский эскиз систематики морфо-функциональных типов органопрофилей. Авторы полагают, что каждому органопрофилю соответствует группа почвенных типов. Поэтому ниже для большинства органопрофилей не приведены ссылки на публикации, за исключением некоторых специфических типов.
Классы органопедогенеза
А. Поверхностно-аккумулятивный органопедогенез (нарастание ПОВ вверх от поверхности минеральной матрицы: подстилка и дернина – торф (горизонты О и Т); далее генезис в сторону формирования органогенных горных пород: бурый уголь – каменный уголь (Шумаков, 1958); предельный случай – это гипер-аккумуляция мощной подстилки в нефелогилее, высокогорных туманных лесах тропиков и экваториальной зоны (Фридланд, 1964; Чертов, 1990).
Б. Профильно-аккумулятивный органопедогенез: накопление ПОВ в гумусово-аккумулятивных минеральных горизонтах ОА, Аh, АЕ и миграция ниже по профилю во всех разновидностях горизонтов Е, В, ВС и С.
Типы органопрофилей в классе Б:
- недифференцированный;
- аккумулятивный (монотонный, горизонты А, В, С);
- гипер-аккумулятивный – чернозём;
- агро-аккумулятивный (все профили с окультуренным Ар, а также российские и немецкие «плаггены» с Ар мощностью до 1 м (Григорьев, 1980; Kalinina et al., 2009);
- элювиальный и иллювиально-элювиальный (АЕ. Е, ЕВ, В);
- альфегумусовый (А, Е, Вfh, B);
- гипер-элювиальный: Ah с интенсивной миграцией ПОВ во влажных тропических лесах (Фридланд, 1964; Чертов, 1985);
- сложный со вторым гумусовым горизонтом (Прокашев, 1999; и др.) или с погребенными профилями;
- аллювиальный;
- вулканический;
- мерзлотный;
- глеевый (Ag, G);
- субаквальный (мелководья, пойменные иловатые топи);
- агросубаквальный (рисовые чеки и др.);
- первичный формирующийся микро-органопрофиль (Абакумов, 2011; Томашунас, Абакумов, 2014; Lessovaia et al., 2008);
- естественно-нарушенный перемешанный, где в наличии имеется ПОВ: эоловый, оползневый, погребенный, ветровально-почвенный и т.д. – с подтипами (Карпачевский и др., 1984; Васенев, Таргульян, 1995; Бобровский, 2010);
- техногенный на разных субстратах, где в наличии имеется ПОВ (Ужегова, Махонина, 1984; Алексеев, 1990; Андроханов и др., 2000; Махонина, 2004; Reintam et al., 2002).
При необходимости должна фиксироваться мощность органопрофиля: маломощный до 30 см, мощный свыше 100 см (средняя категория не обозначается). Приведенный эскиз отражает авторские представления и может служить лишь начальным пунктом в создании обстоятельной систематики органопрофилей почв с учетом существующих концепций и классификаций.
Создание классификационной категории «органопрофиль» будет оправдано в почвенной таксономии и, безусловно, их дальнейшее развитие будет полезным в теоретических работах по эволюции почв. Однако, несмотря на длительное существование этого понятия (Гришина, Орлов, 1978), наблюдается его умеренное использование в почвенно-генетических изысканиях, нередко с редуцированием этой концепции до форм гумуса (Миньковский, Шоба, 1994, 1995; Томашунас, Абакумов, 2014; Бахмет, 2015).
Высказанные положения об органопедогенезе могут быть представлены в виде графической модели процессов педогенеза (рис. 2), которая показывает место органопедогенеза в структурной организации почвенной системы по представлениям авторов.
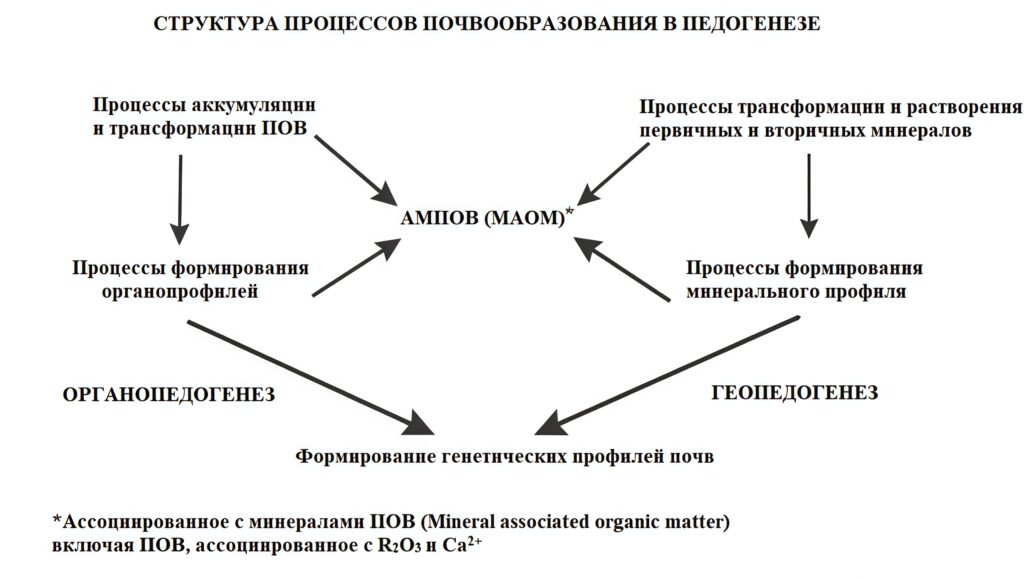
Рисунок 2. Общая структура педогенеза на уровне процессов почвообразования с отражением места органопедогенеза. «Процессы аккумуляции и трансформации ПОВ» относятся к формам гумуса и типам подстилок.
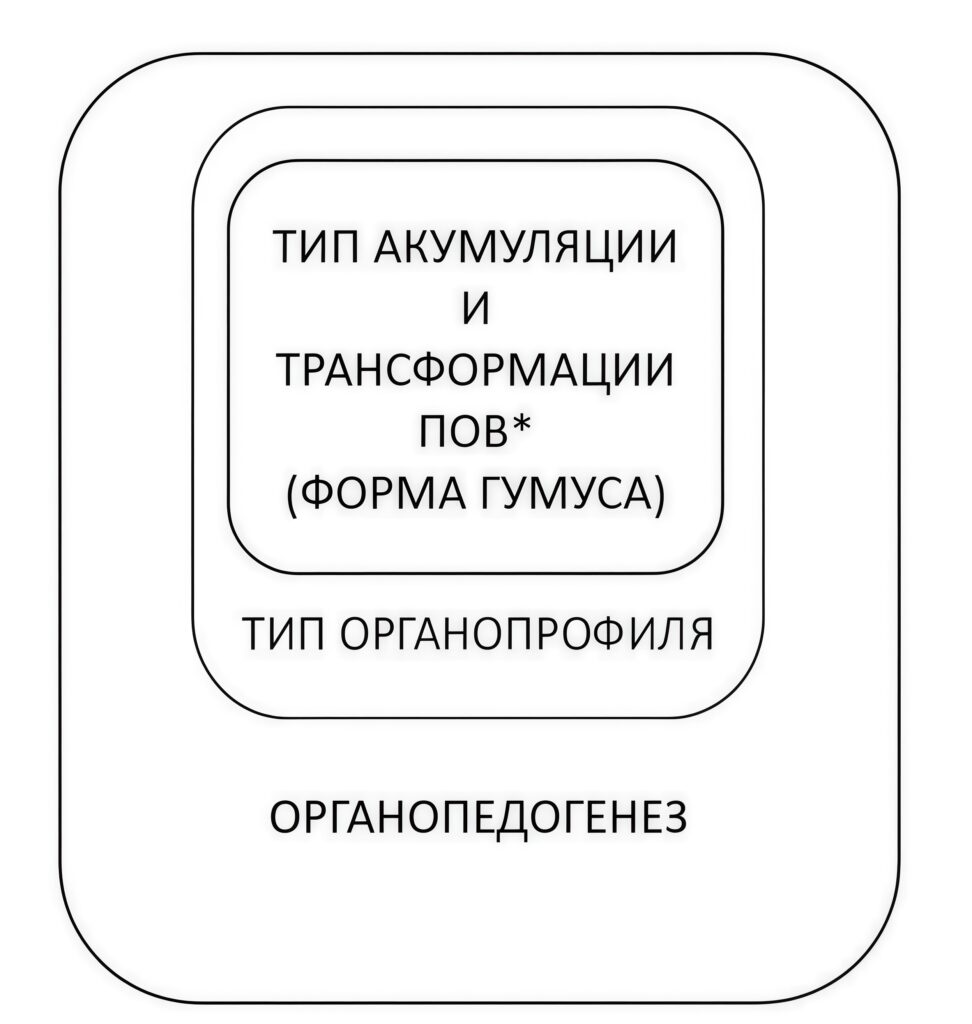
Рисунок 3. Таксономическое соподчинение компонентов органопедогенеза («русская матрёшка»); *ПОВ — почвенное органическое вещество
Так, педогенез в целом охватывает все процессы почвообразования, и его иерархическая дифференциация на органопедогенез и геопедогенез (процессы, затрагивающие только минеральную матрицу: термин предложен авторами) вместо единого почвенного морфогенеза (Шоба, 1988) представляется закономерной. На основном иерархическом уровне логично выделение категории «Типы органопрофиля» и «Типы минерального профиля», а ниже «Формирование типов аккумуляции и трансформации ПОВ», имея также ввиду сложные процессы миграции ПОВ и формирования органо-минеральных комплексов разного генезиса в профиле почвы с выходом в биогеохимический круговорот на ландшафтном, региональном и глобальном уровнях.
С точки зрения организации педогенеза от частного к общему будет полезна другая форма представления в виде «русской матрёшки» (рис. 3), позволяющая понять соподчиненность структурных компонентов общего педогенеза с учетом роли формирования ПОВ. В такой трактовке типы подстилок и других органогенных горизонтов автоморфных позиций логично вписываются в общую структуру органопедогенеза.
Скорости органопедогенеза
Рисунок 3 может быть использован и для демонстрации скорости формирования компонентов органопедогенеза. Тип аккумуляции/трансформации ПОВ и формы гумуса – динамичная структура, включающая наиболее быстро формирующиеся подгоризонты подстилки — подгоризонт опада OL и ферментативный подгоризонт OF со скоростями образования от 1 до 100 лет и гумифицированный подгоризонт подстилки ОH плюс горизонт Аh с временами морфогенеза от 10 до 300 лет. Эти эдафические компоненты, ответственные за биотический круговорот С, N, Р и других элементов-биофилов, обстоятельно изучены по отдельности (Миньковский, Шоба, 1994; Богатырев и др., 2004; Лукина, Никонов, 1996, 1998; Лукина и др., 2019; Солодовников, 2025). Именно эти аккумулятивные почвенные горизонты (topsoil) в наибольшей степени определяют эффективное реализованное плодородие почв в естественных, особенно лесных, экосистемах (Чертов, 1981, 1985; Лукина и др., 2019). При этом необходимо подчеркнуть, что в почвах бореальных лесов, преобладающих в России, подстилка, как правило, довольно мощная (больше 10 см, если не подвержена недавним пожарам), представлена всеми подгоризонтами. Это обусловлено небольшой скоростью переработки опада из-за низкого качества опада (широкое отношение C/N) и низкого уровня биоразнообразия, небольшой численности и биомассы почвенных сапрофагов. Горизонт Аh в этих почвах далеко не всегда выражен, особенно на каменистых песчаных моренах. В хвойно-широколиственных и широколиственных лесах мощность подстилки, напротив, небольшая (1-2 см), подгоризонты подстилки выражены очень неотчетливо из-за активного перемешивания и переработки почвенными сапрофагами. Подгоризонт опада высокого качества с узким отношением C/N может быть выражен осенью, если сразу полностью не переработан, при этом выражен горизонт Аh и также идентифицируется переходный слой между OH и Аh.
Таким образом, качество опада играет существенную роль в формировании типов аккумуляции и трансформации ПОВ (форм гумуса). Л. Г. Богатырев и Т. В. Фомина (1991) при расчете эллювиально-аккумулятивных коэффициентов по А. А. Роде (1984) принимали условно подгоризонт опада (О1) за «материнскую породу». На наш взгляд, опад действительно можно рассматривать как вторую материнскую породу, поскольку качество опада, так же как состав почвообразующей породы, играет большую роль в почвообразовании.
В отличие от форм гумуса, органопрофиль – это уже почвенно-генетическая категория с временами формирования от 100 лет до тысячелетий с главной ролью в развитии полного почвенного профиля с трансформацией и растворением минералов за счет энергии минерализуемого биотой ПОВ и с выходом органических и минеральных веществ из малого биотического круговорота в большие биогеохимические циклы. При этом, в одном и том же органопрофиле может формироваться несколько форм гумуса. Фактически форма гумуса вложена в органопрофиль, как самая верхняя аккумулятивная часть, стартер и драйвер процессов формирования ПОВ. Но объединение формы гумуса с органопрофилем в одну категорию будет концептуально некорректно в силу различий их роли и скоростей формирования. С другой стороны, вполне возможно, что типы органопрофилей послужат основой для создания общих типов почвенного морфогенеза в контексте представлений С. А. Шобы (1988). В геопедогенезе периоды формирования полного минерального профиля (Таргульян, 2019) в разы длительнее периодов формирования органопрофиля в органопедогенезе.
Скорость восстановления формы гумуса и органопрофиля при их физическом нарушении (например, при ветровалах) достаточно высока под пологом растительности в зависимости от качества и мощности потока опада и сохранности фрагментов аккумулятивных горизонтов (Карпачевский и др., 1984; Разумовский, 1999; Васенев, 2008). Однако она будет медленной при исходном отсутствии растительности на экспонированной материнской породе (горизонт С), как и в ходе первичной экогенетической сукцессии (Разумовский, 1999; Abakumov, 2001). При техногенном нарушении восстановление формы гумуса и органопрофиля происходит в зависимости от мощности и режима нарушения (загрязнение, рекреация), а также свойств техногенного субстрата (Алексеев, 1990; Андроханов и др., 2000). Рассмотренные выше скорости процессов формирования и восстановления форм гумуса и органопрофилей демонстрируют влияние классического фактора почвообразования «время» на процессы органопедогенеза. Более того, эти параметры позволяют также оценить максимальное время реакции компонентов органопедогенеза на изменение природных условий, более всего на современное изменение климата. В настоящее время более точные оценки этих процессов при отсутствии экспериментальных данных и хроно-рядов возможны с использованием существующих почвенных и экосистемных моделей (например, Шанин и др., 2022; Komarov et al., 2017).
Включение компонентов органопедогенеза в классификацию почв
Как отмечалось выше, еще в первой половине ХХ в. И. В. Тюрин и В. В. Пономарёва (1940) обозначали формы гумуса прилагательным к слову «почва» (например, муллевая почва, грубогумусная почва). Н. Л. Благовидов и Г. Л. Бурков (1959) рекомендовали и использовали этот формат в классификации лесных почв (например, модергумусная среднеподзолистая легкосуглинистая…). С тех пор этот шаблон наименования почв по формам гумуса использовался в почвенных и лесотипологических работах на Северо-Западе Европейской России (Чертов, 1966, 1981; Чертов, Дыренков, 1973; Чертов и др., 1978; Дыренков, 1984; Федорчук и др., 2005; Чертов, Надпорожская, 2018), а формы/типы гумуса были подробно описаны даже с полевым дихотомическим определителем (Чертов, 1974).
Все высказанные положения определяют необходимость классификационного отражения роли органопедогенеза в почвообразовании. Здесь также имеется определённый положительный опыт (Чертов, 1974, 1981) и, более того, предложен методический подход по расширению российской классификации почв для включения в неё форм гумуса как типов аккумуляции и трансформации ПОВ (Чертов, 2025).
В соответствии с этим подходом, при включении компонентов органопедогенеза в актуальную российскую почвенную классификацию (Классификация и диагностика …, 2004; Полевой определитель …, 2008) типы органопрофилей могут занять место неясно определенного подтипа почв, а типы аккумуляции и трансформации ПОВ могут быть интегрированы ниже, поскольку формы гумуса представляют собою наиболее динамичную морфологическую структуру почвы. Для этой таксономической единицы предлагается использовать существующий в российской классификации уровень «род» в соответствии с предложением М. И. Герасимовой (Герасимова, 2018; Хитров, Герасимова, 2022), который был введен для отражения как роли почвообразующих пород и загрязнения, так и для временных и механизменно-процессных категорий. Особенностью интеграции в него типов аккумуляции и трансформации ПОВ (форм гумуса) являются: а) некое смешение таксономического порядка (признаки – типы) и б) обязательность его введения для всех почвенных таксонов ниже уровня типа (для которых формы гумуса уже существуют). Такая таксономическая архитектура не нарушает общей композиции генетической классификации почв и ни в коей мере не претендует на его замену, не требуя никаких структурных перестроек в существующей классификации (не вместо генетической, а совместно с генетической).
В итоге получается сочетание эдафологической классификации ПОВ с генетической для всей почвы, принимая во внимание, что гумус нижней части почвенного профиля более стабилен и отражает как историю почвы, так и современные почвенные процессы. В этом случае почвенная карта трансформируется из строгого классического портрета в живую динамичную картину, которая потребует периодического обновления (в лесных экосистемах в интервале 10 – 100 лет) для отслеживания органопедогенеза под влиянием меняющихся природных и антропогенных факторов.
Реализация высказанных предложений по сочетанию типологических структур органопедогенеза с генетическими таксонами потребует содержательного анализа существующей таксономии для понятной и приемлемой интеграции этих компонентов, а также обстоятельной проработки систематики типов органопрофилей в развитие предложенного выше «эскиза». При всей простоте алгоритма объединения здесь могут встретиться некоторые сложности. Например, может возникнуть ситуация неопределенности, когда непонятно какой тип органопрофиля или формы гумуса представлен у исследуемой почвы (например, у такыра или солончака). В этом случае следует просто опустить указание органопрофиля и формы гумуса, как наиболее логичное и простое решение, при этом важно сохранить описание ПОВ для анализа и заключения о том, является ли это новым органопрофилем или формой гумуса либо это некий вариант уже имеющихся таксонов. Следует также иметь ввиду, что в российской генетической классификации генетические названия почв и формы гумуса совпадают на гидроморфных органогенных почвах и, безусловно, здесь приоритет должен отдаваться генетическому определению.
Предложенная концепция органопедогенеза и процедура её включения в почвенную таксономию могут вызвать различные возражения. Однако это не меняет постулата о необходимости обязательного и достаточно детального отражения морфологии ПОВ в почвенной классификации, что важно как с общетеоретической биосферной, так и с «прикладной» эдафической позиций. До сих пор прогресса в этой области не наблюдалось и это даже не обсуждается (Лебедева и др., 2000; Тонконогов и др., 2005).
Тем не менее, в лесном почвоведении и лесоведении формы гумуса уже давно включены в методические руководства по картографированию лесных почв, но, к сожалению, пока не используются в России, хотя используются в большом количестве зарубежных стран (Малышева и др., 2022). В последнее время формы гумуса также внедряются в методические материалы важнейшего инновационного проекта государственного значения (ВИП ГЗ) «Единая национальная система мониторинга климатически активных веществ» и государственной инвентаризации лесов России».
ЗАКЛЮЧЕНИЕ
Предложенная нами концепция органопедогенеза, во-первых, может расширить теоретический фундамент современного почвоведения с акцентом на важнейшую роль биологического фактора в функционировании педосферы, в том числе в условиях современных изменений климата. Это не только влияние растительности, но в значительной степени и всей почвенной биоты. Во-вторых, есть надежда, что она сможет объединить генетическую и эдафическую ветви почвоведения в представление о целостной быстро эволюционирующей почвенной системе, как с теоретической биосферной позиции, так и, особенно, с точки зрения практического использования при решении задач оптимизации природопользования в условиях динамичных изменений окружающей среды и возрастания антропогенного прессинга на биосферу планеты.
Авторы выражают благодарность уважаемым рецензентам за высказанные ими ценные замечания по содержанию этой работы, что позволило существенно улучшить изложение высказанных положений.
ФИНАНСИРОВАНИЕ
Работа выполнена за счет средств государственного задания по теме «Биоразнообразие и экосистемные функции лесов» (Регистрационный номер НИОКТР 124013000750-1).
СПИСОК ЛИТЕРАТУРЫ
Абакумов Е. В. Хронология онтогенеза первичных почв: обзор проблемы // Вестник Санкт-Петербургского университета. Серия 3. 2011. Вып. 3. С. 114–120.
Алексеев В. А. Лесные экосистемы и атмосферное загрязнение. Ленинград: Наука, 1990. 200 с.
Андроханов В. А., Овсянникова С. В., Курачев В. М. Техноземы: свойства, режимы, функционирование. Новосибирск: Наука, 2000. 200 с.
Бахмет О. Н. Структурно-функциональная организация органопрофилей почв лесных экосистем Северо-Запада России. Автореф. дисс. докт. биол. наук. Петрозаводск, 2015. 49 с.
Биоразнообразие и функционирование лесных экосистем / Под ред. Н. В. Лукиной. Москва: КМК, 2021. 327 с.
Благовидов Н. Л., Бурков Г. Л. Методические указания к производству почвенных исследований и характеристике условий местообитания леса. Ленинград: ЛТА, 1959. 30 с.
Бобровский М. В. Лесные почвы европейской России. Биотические и антропогенные факторы формирования. М.: КМК, 2010. 392 с.
Богатырев Л. Г., Демин В. В., Матышак Г. В., Сапожникова В. А. О некоторых теоретических аспектах исследования лесных подстилок. Лесоведение. 2004. № 4. С. 17–29.
Богатырев Л. Г., Фомина Т. В. Характеристика подстилок сосняков Присурского лесного массива // Вестник Московского университета. Серия 17. 1991. № 3. С. 28–39.
Будыко М. И. Эволюция биосферы. Л.: Гидрометеоиздат, 1984. 488 с.
Ваксман З. А. Гумус: Происхождение, химический состав и значение его в природе. Москва: Сельхозгиз, 1937. 471 с.
Васенев И. И. Почвенные сукцессии. М.: ЛКИ, 2008. 400 с.
Васенев И. П., Таргульян В. О. Ветровал и таежное почвообразование. М: Наука. 1995. 247 с.
Вернадский В. И. Биосфера. Ленинград: Науч. хим.-технол. изд-во. 1926. 147 с.
Вернадский В. И. Живое вещество и биосфера. М.: Наука, 1994. 671 с.
Вильямс В. Р. Почвоведение. Избранные сочинения. Москва: Издательство «Юрайт», 2025. 344 с.
Герасимова М. И. Нужен ли уровень рода в классификации почв России? // Бюллетень Почвенного института им. В. В. Докучаева. 2018. Вып. 95. С. 90–98. DOI: 10.19047/0136-1694-2018-95-90-98
Глазовская М. А. Геохимия природных и техногенных ландшафтов СССР. М.: Высшая школа, 1988. 327 с.
Горшков В. Г. Физические и биологические основы устойчивости жизни. М., 1995. 470 с.
Григорьев Г. И. Диагностические показатели степени окультуренности дерново-подзолистых, светло-серых и серых лесных суглинистых почв // Теоретические основы и методы определения оптимальных параметров свойств почв. М., 1980. С. 21–29.
Гришина Л. А. Гумусообразование и гумусное состояние почв. М.: Изд-во МГУ, 1986. 244 с.
Гришина Л. А., Орлов Д. С. Система показателей гумусного состояния почв // Проблемы почвоведения (Советские почвоведы к II Международному конгрессу почвоведов в Канаде, 1978 г.). М.: 1978. С. 42–47.
Данилов Д. А., Яковлев А. А., Зайцев Д. А., Иванов А. А. Изменения в постагрогенных почвах в ходе восстановления древесной растительности в условиях юго-запада Ленинградской области. Известия СПбЛТА. 2024. № 251. С. 97–122. DOI: 10.21266/2079-4304.2024.251.97-122
Добровольский Г. В. Педосфера – оболочка жизни планеты Земля. Биосфера. 2009. Т. 1. № 1. С. 6–14.
Добровольский Г. В., Карпачевский Л. О., Криксунов Е. А. Геосферы и педосфера. М.: ГЕОС, 2010.190 с.
Докучаев В. В. Русский чернозём: Отчёт Вольному экономическому обществу. СПб.: Тип. Деклерона и Евдокимова, 1883. 376 c.
Дыренков С. А. Структура и динамика таежных ельников. Л.: Наука, 1984. 176 с.
Дюшофур Ф. Основы почвоведения. Эволюция почв. Москва: Прогресс, 1979. 592 с.
Карпачевский Л. О. Лес и лесные почвы. Москва: Лесная промышленность, 1981. 264 с.
Карпачевский Л. О., Строганова M. П., Таргульян В. О., Васенев И. И., Гончарук Н. Ю. Почвенные микросукцессии в климаксных ельниках южной тайги // История развития почв СССР в голоцене. Пущино: АН СССР, 1984. С. 68–69.
Классификация и диагностика почв России / Авторы и составители: Л. Л. Шишов, В. Д. Тонконогов, И. И. Лебедева, М. И. Герасимова. Смоленск: Ойкумена, 2004. 342 с.
Ковда В. А. Биогеохимия почвенного покрова. М., 1985. 493 с.
Костычев П. А. Почвы черноземной области России, их происхождение, состав и свойства. СПб.: Изд. Девриена, 1886. 231 с.
Крюденер А. А. Основы классификации типов насаждений. Материалы по изучению русского леса. Петроград, 1916. 190 с.
Лебедева H. H., Тонконогов В. Д., Герасимова М. И. Опыт разработки факторной классификации почв // Почвоведение. 2000. № 2. С. 148–157.
Лукина Н. В. Влияние современных изменений климата на лесные почвы // Тезисы доклада. Семинар Института глобального климата и экологии имени академика Ю. А. Израэля 30 апреля 2025 г. ИГКЭ, 2025. URL: http://downloads.igce.ru/seminars2/Lukina_N_V_Abstract_IGCE_30042025.pdf (дата обращения 01.07.2025)
Лукина Н. В., Барталев С. А., Гераськина А. П., Плотникова А. С., Горнов А. В., Ершов, Д. В., … & Ручинская Е. В. Роль старовозрастных лесов в аккумуляции и хранении углерода // Известия Российской академии наук. Серия географическая. 2023. Т. 87. №. 4. С. 536–557.
Лукина Н. В., Гераськина А. П., Кузнецова А. И., Смирнов В. Э., Горнов А. В., Шевченко Н. Е., Тихоновa Е. В. Функциональная классификация лесов: актуальность и подходы к разработке // Лесоведение. 2021. № 6. С. 566–580.
Лукина Н. В., Кузнецова А. И., Гераськина А. П., Смирнов В. Э., Иванова В. Н., Тебенькова Д. Н., Горнов А. В., Шевченко Н. Е., Тихонова Е. В. Неучтенные факторы, определяющие запасы углерода в лесных почвах // Метеорология и гидрология. 2022. № 10. С. 92–110. DOI: 10.52002/0130-2906-2022-10-92-110
Лукина Н. В., Никонов В. В. Биогеохимические циклы в лесах Севера в условиях аэротехногенного загрязнения. Апатиты: Кольский научный центр РАН, 1996. 192 с.
Лукина Н. В., Никонов В. В. Питательный режим лесов северной тайги: природные и техногенные аспекты. Апатиты: Кольский научный центр РАН, 1998. 315 с.
Лукина Н. В., Орлова М. А., Бахмет О. Н., Тихонова Е. В., Тебенькова Д. Н., Казакова А. И., Крышень А. М., Горнов А.В., Смирнов В. Э., Шашков М. П., Ершов В. В., Князева С. В. Влияние растительности на характеристики лесных почв Республики Карелия // Почвоведение. 2019. № 7. C. 827–842.
Лукина Н. В., Орлова М. А., Исаева Л. Г. Плодородие лесных почв как основа взаимосвязи почва-растительность // Лесоведение. 2010. № 5. С. 45–56.
Лукина Н. В., Полянская Л. М., Орлова М. А. Питательный режим почв северотаежных лесов. Москва: Наука, 2008. 341 с.
Малышева Н. В., Филипчук А. Н., Золина Т. А., Кинигопуло П. С., Шалимова Е. М., Попик С. А., Сильнягина Г. В. Анализ зарубежного опыта национальных инвентаризаций лесов: методы, выборка, результаты и международная статистика // Лесохозяйственная информация. 2022. № 2. С. 90–132. DOI: 10.24419/LHI.2304-3083.2022.2.08
Махонина Г. И. Начальные процессы почвообразования в техногенных экосистемах Урала. Автореф. дис. … д-ра биол. наук. Томск, 2004. 38 с.
Миньковский Г. М., Шоба С. А. Морфология и классификация органопрофилей почв южной тайги // Почвоведение. 1994. № 9. С 90–101.
Миньковский Г. М., Шоба С. А. Морфоструктурные подходы к типизации органопрофилей почв // Почвоведение. 1995. № 10. С. 1271–1283.
Мировая реферативная база почвенных ресурсов. Международная система почвенной классификации для диагностики почв и составления легенд почвенных карт: перевод с англ. / Под ред. М. И. Герасимовой и П. В. Красильникова. 4-е издание. Международный союз наук о почве, Москва, 2024. М.: МАКС Пресс, 2024. 248 с.
Одум Ю. Экология. Т. 2. М.: Мир, 1986. 376 с.
Орлов Д. С., Бирюкова O. Н., Суханова Н. И. Органическое вещество почв Российской Федерации. М.: Наука. 1996. 256 с.
Перельман А. И. Геохимия ландшафта. М.: Высшая школа, 1975. 341 с.
Полевой определитель почв. М.: Почвенный ин-т им. В.В. Докучаева, 2008. 182 с.
Пономарева В. В. Теория подзолообразовательного процесса. M.-JI.: Наука, 1964. 379 с.
Прокашев A. M. Почвы со сложным органопрофилем юга Кировской области. Киров, 1999. 176 с.
Разумовский С. М. Избранные труды. Москва: KMK, 1999. 560 с.
Роде А. А. Генезис почв и современные факторы почвообразования. М.: АН СССР, 1984. 256 с.
Родин Л. Е., Базилевич Н. И. Динамика органического вещества и биологический круговорот зольных элементов и азота в основных типах растительности земного шара. Акад. наук СССР. Ботан. ин-т им. В. Л. Комарова. Москва; Ленинград: Наука. 1965. 253 с.
Семенов В. М., Когут Б. М. Почвенное органическое вещество. М.: ГЕОС, 2015. 233 с.
Семенов В. М., Лебедева Т. Н., Соколов Д. А., Зинякова Н. Б., Лопес де Гереню В. О., Семенов М. В. Измерение почвенных пулов органического углерода, выделенных био-физико-химическими способами фракционирования // Почвоведение. 2023. № 9. С. 1155–1172. DOI: 10.31857/S0032180X23600427
Сибирцев Н. М. Почвоведение. СПб.: Изд. Скворцова, 1899. 360 с.
Смирнова О. В. Популяционная организация биоценотического покрова лесных ландшафтов // Успехи современной биологии. 1998. Т. 118. № 2. С. 148–165.
Смирнова О. В., Торопова Н. А. Потенциальная растительность и потенциальный экосистемный покров // Успехи современной биологии. 2016. Т. 136. № 2. С. 199–211.
Смирнова О. В., Торопова Н. А. Сукцессия и климакс как экосистемный процесс // Успехи современной биологии. 2008. Т. 128. № 2. С. 129–144.
Соколов И. А. Почвообразование и экзогенез. М., 1997. 244 с.
Солодовников А. Н. Модели начального разложения хвои сосны обыкновенной в среднетаежных условиях Восточной Фенноскандии // Бюллетень Почвенного института имени В. В. Докучаева. 2025. Вып. 122. С. 41–61. DOI: 10.19047/0136-1694-2025-122-41-61
Сукачев В. Н., Дылис Н. В. Основы лесной биогеоценологии. М. Наука, 1964. 573 с.
Таргульян В. О. Теория педогенеза и эволюции почв. М.: ГЕОС, 2019. 295 с.
Томашунас В. М., Абакумов Е. В. Органопрофили почв полуостровов Ямал и Гыдан // Международная научн. конф. «Экология и биология почв». Ростов-на-Дону, 2014. С. 364–365.
Тонконогов В. Д., Лебедева И. И., Герасимова М. И., Красильников П. В., Дубровина И. А. Корреляция почвенных классификаций. Петрозаводск: КарНЦ РАН, 2005. 52 с.
Трофимов С. С., Таранов С. А., Наплекова Н. Н., Фаткулин Ф. А. Гумусообразование в техногенных экосистемах. Новосибирск: Наука, 1986. 126 с.
Тюрин И. В., Пономарёва В. В. Материалы по изучению гумуса лесных почв // Труды Ленинградской лесотехнической академии. 1940. Вып. 56. С. 3–49.
Ужегова И. А., Махонина Г. И. Начальные процессы почвообразования на отвалах Первоуральского месторождения железных руд // Почвоведение. 1984. № 11. С. 14−21.
Федорчук В. Н., Нешатаев В. Ю., Кузнецова М. Л. Лесные экосистемы северо-западных районов России. Типология, динамика, хозяйственные особенности / Отв. ред. О. Г. Чертов. СПб: Санкт-Петербургский НИИ лесн. хоз-ва. 2005. 381 с.
Фокин А. Д. Динамическая характеристика гумусового профиля подзолистой почвы // Известия Тимирязевской сельскохозяйственной академии. 1975. № 4. С. 80–88.
Фридланд В. М. Почвы и коры выветривания влажных тропиков. АН СССР. Москва: Наука, 1964. 312 с.
Хитров Н. Б., Герасимова М. И. Предлагаемые изменения в классификации почв России: диагностические признаки и почвообразующие породы // Почвоведение. 2022. № 1. С. 3–14. DOI: 10.31857/S0032180X22010087
Чертов О. Г. Об экологических функциях и эволюции почв. Вестник ЛГУ. 1990. Сер. 3. № 2(10). C. 75–81.
Чертов О. Г. Определение типов гумуса лесных почв. Методические указания. Л.: Лен. НИИ лесн. хоз-ва, 1974. 16 с.
Чертов О. Г. Экология лесных земель. Почвенно-экологическое исследование лесных местообитаний. Ленинград: Наука, 1981. 192 с.
Чертов О. Г. Экотопы тропического дождевого леса. Ленинград: Наука, 1985. 47 с.
Чертов О. Г. О смысловой структуре наименований таксонов почвенных классификаций // Вопросы лесной науки. 2025. Т. 8. № 1. Статья № 163. DOI: 10.31509/2658-607x-202581-163
Чертов О. Г., Дыренков С. А. О классификации лесных биогеоценозов Карельского перешейка // Труды Петрозаводской лесной опытной стации. Л.: ЛенНИИЛХ, 1973. Т. 2. С. 74–80.
Чертов О. Г., Мельницкая Г. Б., Ламов А. К., Берг И. Е. Опыт работ по лесной типологии на основе картирования почв и местообитаний // Сборник статей по лесоводству. Ленинград: Лен. НИИ лесн. хоз-ва. 1974. № 17. С. 24–47.
Чертов О. Г., Надпорожская М. А. Формы гумуса лесных почв: концепции и классификации // Почвоведение. 2018. № 10. С. 1202–1214.
Чертов О. Г., Надпорожская М. А., Палёнова М. М., Припутина И. В. Эдафология в структуре почвоведения и экосистемной экологии // Russian Journal of Ecosystem Ecology. 2018. Vol. 3 No. 3. DOI: 10.21685/2500-0578-2018-3-2
Чертов О. Г., Разумовский С. М. Об экологической направленности процессов развития почв // Журнал общей биологии. 1980. Т. 62. № 3. С. 386–396.
Чертов О. Г., Филиппов Г. В., Мельницкая Г. Б. Крупномасштабное картирование почв при лесоустройстве. Методические указания. Ленинград: Лен. НИИ лесн. хоз-ва, 1978. 52 с.
Чуков С. Н. Применение метода ядерного магнитного резонанса к изучению органического вещества почв // Вестник Санкт-Петербургского университета. Серия биологическая. 1997. Вып. 4. С. 50–59.
Шанин В. Н., Фролов П. В., Припутина И. В., Чертов О. Г., Быховец С. С., Зубкова Е. В., Портнов А. М., Фролова Г. Г., Стаменов М. Н., Грабарник П. Я. Моделирование динамики лесных экосистем с учётом их структурной неоднородности на разных функциональных и пространственных уровнях // Вопросы лесной науки. 2022. Т. 6 № 3. DOI: 10.31509/2658-607x-202252-112
Шоба С. А. Морфогенез почв лесной зоны. Автореферат. дис. докт. биол. наук. М., 1988. 48 с.
Шумаков B. C. Принципы классификации, номенклатуры и картирования лесных подстилок // Сборник работ по лесному хозяйству. Вып. 35. М.–Л.: Гослесбумиздат, 1958. С. 199–216.
Эволюция почв и почвенного покрова. Теория, разнообразие природной эволюции и антропогенных трансформаций почв / отв. ред. В. Н. Кудеяров, И. В. Иванов. М.: ГЕОС, 2015. 928 с.
Эволюция, функционирование и экологическая роль почв как компонентов биосферы / отв. ред. А. О. Алексеев. Москва: КМК, 2020. 290 с.
Ярилов А. А. Педология как самостоятельная естественнонаучная дисциплина о земле. Юрьев (Тарту): Изд. Юрьевского ун-та, 1904–1905. Часть 1. 480 с.; Часть 2. 244 с.
Abakumov E. V. Soils of mining areas – natural model of soil formation // Proceed. 1st Intern. Conf. on Soils and Archaeology. Szazhalombatta, Hungary, 2001. Р. 163−165.
Broll G., Brauckmann H.-J., Overesch M., Junge B., Erber C., Milbert, G., Baize D., Nachtergaele F. Topsoil characterization – Recommendations for revision and expansion of the FAO-Draft (1998) with emphasis on humus forms and biological features // Journal of Plant Nutrition and Soil Science. 2006. Vol. 169. No. 3. P. 453–461.
Chertov O. G., Nadporozhskaya M. A. Development and application of humus form concept for soil classification, mapping and dynamic modelling in Russia // Applied Soil Ecology. 2018. Vol. 122. P. 420–423. DOI: 10.1016/j.apsoil.2017.04.006
Chertov O., Gorbushina A., Deventer B. A model for microcolonial fungi growth on rock surfaces // Ecological Modelling. 2004. Vol. 177. No. 3-4. P. 415–426. DOI: 10.1016/j.ecolmodel.2004.02.011
Duchaufour Ph. Precis de Pedologie. 2nd edition, Paris: Masson, 1965. 482 p.
Emmer I. M. Humus form and soil development during a primary succession of monoculture Pinus sylvestris forests on poor sandy substrates. Amsterdam, 1995. 135 p.
Green R. N., Trowbridge R. L., Klinka K. Towards a taxonomic classification of humus forms // Forest Science. 1993. Vol. 39. No. 1. P. 1–56.
Humusica – Terrestrial Natural Humipedons / A. Zanella, J. Ascher-Jenull (eds.) // Applied Soil Ecology. 2018. Vol. 122. Part I, II, III.
IUSS Working Group WRB. World Reference Base for Soil Resources. International soil classification system for naming soils and creating legends for soil maps. 4th edition. International Union of Soil Sciences (IUSS), Vienna, Austria. 2022. DOI: 10.29003/m4174.978-5-317-07235-3.
Kalinina O., Chertov O., Nadporozhskaya M., Giani L. Properties of soil organic matter of Plaggic Anthrosols from Northwest Germany, Northwest and North Russia // Archives of Agronomy and Soil Science. 2009. Vol. 55. No. 5. P. 477–492. DOI: 10.1080/03650340802637733
Klinka K., Krestov P., Fons J., Chourmouzis C. Towards a taxonomic classification of humus forms: Third approximation // Scientia Silvia. 1997. No. 9. P. 1–5.
Kõlli R., Köster T. Interrelationships of humus cover (pro humus form) with soil cover and plant cover: humus form as transitional space between soil and plant // Applied Soil Ecology. 2018. Vol. 123. P. 451–454.
Komarov A., Chertov O., Bykhovets S., Shaw C., Nadporozhskaya M., Frolov P., Shashkov M., Shanin V., Grabarnik P., Priputina I., Zubkova E. Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing // Ecological Modelling. 2017. Vol. 345. P. 113–124. DOI: 10.1016/j.ecolmodel.2016.08.007
Kubiena W. L. The soils of Europe. London: Thomas Murby Co., 1953. 318 p.
Lehmann J., Kleber M. The Contentious Nature of Soil Organic Matter // Nature. 2015. Vol. 528. P. 60–68. DOI: 10.1038/nature16069
Lessovaia S., Chertov O., Goryachkin S. Specificity of pedogenesis in shallow soils on massive rocks of East Fennoscandia // Agricultural Sciences/Zemès ukio Mokslai. 2008. Vol. 12. No. 3. P. 80–86.
Lukina N. V., Tikhonova E. V., Danilova M. A., Bakhmet O. N., Kryshen A. M., …, & Zukert N. V. Associations between forest vegetation and the fertility of soil organic horizons in northwestern Russia // Forest Ecosystems. 2019. P. 1–19. doi: 10.1186/s40663-019-0190-2
Müller P. E. Studien über die natürlichen Humusformen und deren Entwicklung auf Vegetation und Boden. Berlin, 1887. 324 p.
Nadporozhskaya M. A., Mohren G. M. J., Chertov O. G., Komarov A. S., Mikhailov A. V. Dynamics of soil organic matter in primary and secondary forest succession on sandy soils in The Netherlands: An application of the ROMUL model // Ecological Modelling. 2006. Vol. 190. P. 399–418. DOI: 10.1016/j.ecolmodel.2005.03.025
Piccolo A., Spaccini R., Savy D., Drosos M., Cozzolino V. The Soil Humeome: Chemical structure, functions and technological perspectives. In: Sustainable Agrochemistry / Vaz Jr. Silvio (ed.). Springer, Cham, 2019. DOI: 10.1007/978-3-030-17891-8_7
Ponge J.-F. Humus forms in terrestrial ecosystems: a framework to biodiversity // Soil Biology and Biochemistry. 2003. Vol. 35. No. 7. P. 935–945. DOI: 10.1016/s0038-0717(03)00149-4.hal-00498465
Reintam L., Elmar K., Rooma I. Development of soil organic matter under pine on quarry detritus of open-cast oil-shale mining // Forest Ecology and Management. 2002. Vol. 171. Р. 191−198.
Wachendorf C., Milbert G., Broll G., Frank T., Graefe U., Beylich A. A concept for a consolidated humus form description – An updated version of the German humus form systematics // International Journal of Plant Biology. 2023. Vol. 14. No. 3. P. 658–686.
Wilde S. A. Forest Soils: Origin, Properties, Relation to Vegetation and Silvicultural Management. New York: Wiley & Sons, 1958. 537 p.
Zanella A., Ponge J.-F., Briones M. J. I. Terrestrial humus systems and forms – Biological activity, space-time dynamics // Applied Soil Ecology. 2018. Vol. 122. Part 1. P. 103–137. DOI: 10.1016/j.apsoil.2017.07.020
Zanella A., Ponge J.-F., Jabiol B., Van Delft B., De Waal R., Katzensteiner K., … & Hager H. Standardized Morpho-Functional Classification of the Planet’s Humipedons // Soil Systems. 2022. Vol. 6. No. 3. Article 59. DOI: 10.3390/soilsystems6030059
Рецензенты: д. б. н., член-корр. РАН Красильников П. С.
д. б. н., профессор Герасимова М. И.
[1] Настоящая работа выполнена по итогам многолетних исследований авторов, поэтому содержит значительное количество их публикаций, а также ссылок на давние работы классиков, что необходимо для освещения предлагаемого подхода. Авторы благодарны редакции и рецензентам за понимание.
[2] Более компактным является термин «органогенез», но он занят в зоологии и медицине как формирование органов у эмбрионов. Кстати, также оказался занятым в зоологии с ХIX в. и термин «педогенез», как один из типов партеногенеза.





