- DOI: 10.31509/2658-607x-202691-187
- УДК 630*182.5:574.36: 630*114(470.21)
Сравнение оценки фитомассы древостоя с применением моделей на примере еловых фитоценозов Мурманской области
![]() © 2026 Н. С. Рябов*, Л. Г. Исаева
© 2026 Н. С. Рябов*, Л. Г. Исаева
Институт проблем промышленной экологии Севера КНЦ РАН
Россия, 184209, Апатиты, Академгородок, 14а
*E-mail: n.ryabov@ksc.ru
Поступила в редакцию: 04.02.2026
После рецензирования: 16.03.2026
Принята к печати: 20.03.2026
Статья посвящена сравнительному анализу эффективности различных регрессионных моделей для оценки фитомассы древостоя в ельниках, сформированных на Al-Fe-гумусовых подзолах на северном пределе распространения древесных растений в Мурманской области. Исследование проводилось на пробных площадях, заложенных в различных позициях ландшафта: автоморфных, транзитных и аккумулятивных. Актуальность работы обусловлена необходимостью получения достоверных данных о запасах фитомассы в лесных экосистемах, поскольку фитомасса – ключевой резервуар углерода таёжных фитоценозов. Неоднородный холмистый и пересеченный рельеф региона определяет необходимость учёта данного фактора для минимизации ошибок в оценках пула фитомассы. Сравнение четырёх моделей показало, что для оценки фитомассы древостоя наиболее применима модель, разработанная J. Repola с соавторами. Так как модель основана на большом массиве данных в сходных природно-климатических условиях, она демонстрирует высокую предсказуемость результатов во всех типах элементарных ландшафтов и является оптимальным компромиссом для оценки пулов фитомассы региона на текущий момент, несмотря на выявленные ограничения при анализе внутренней фракционной структуры фитомассы. Региональная модель, в свою очередь, ограничена учетом исключительно надземной фитомассы и может быть использована для оценки данной части в автоморфных условиях. Обнаружено, что модель В. А. Усольцева пригодна для оценки фитомассы ели, однако выявлено систематическое занижение этого показателя для березы – основной сопутствующей породы в северотаёжных ельниках. Выявлена неэффективность для научно-обоснованного мониторинга модели, которая предложена в методических материалах Минприроды РФ, поскольку отмечены значительные завышения фитомассы березовой части древостоя. Отсутствие надежных аллометрических уравнений для оценки подземной фитомассы (корней) является существенным пробелом, как и выявленные неопределённости в оценках фракционной структуры фитомассы деревьев. Результаты исследования свидетельствуют о необходимости получения экспериментальных данных для уточнения региональных моделей, в том числе с учетом позиций ландшафта. Это позволит минимизировать ошибки при калибровке дистанционных методов зондирования и повысить точность количественного учета роли лесов Мурманской области в глобальном цикле углерода.
Ключевые слова: фитомасса древостоя, модели оценки фитомассы, еловые леса, северная тайга, Мурманская область, Арктика
Ведущая роль лесов в депонировании атмосферного углерода обусловливает важность оценки запасаемой фитомассы в этих экосистемах. Углерод входит в состав парниковых газов, влияющих на наблюдаемые климатические изменения. В настоящее время наблюдается дисбаланс углеродного цикла на планете. По современным оценкам, Российская Федерация является крупнейшей страной – поглотителем парниковых газов (Friedlingstein et al., 2025). На леса бореальной зоны приходится около трети всего углеродного стока наземных экосистем (Pappas et al., 2020). Однако согласно исследованиям Pan et al. (2024), вследствие сокращения площади бореальных лесов снижается поглощение атмосферного углерода этими экосистемами (-36 ± 6%).
На сегодняшний день известно, что малонарушенные старовозрастные лесные биогеоценозы, в которых преобладают естественные процессы, являются уникальными природными комплексами, рефугиумами биоразнообразия (Зайцева и др., 2002; Framstad et al., 2013) и накопителями углерода (Лукина и др., 2023; Luyssaert et al., 2008; Framstad et al., 2013). Относительно недолгая история промышленной заготовки леса в Мурманской области, а также отсутствие традиции у коренного населения практики подсечно-огневого земледелия позволяют рассматривать современные перестойные насаждения как малонарушенные биогеоценозы с естественной пространственной и возрастной структурой (Экологический…, 1999). Согласно лесному плану Мурманской области (Лесной план…, 2019), на долю перестойных хвойных лесов приходится 33.7% всего запаса насаждений (23% лесопокрытой площади). При сравнении с соседними странами – Финляндией, Швецией и Норвегией, где таких лесов, соответственно, только 10, 11 и 15% (Framstad et al., 2013), можно сделать вывод о значительном потенциале заполярных лесных экосистем Северо-Запада РФ в секвестрации углерода. При этом, ввиду более интенсивного использования сосновых лесов, в регионе на долю спелых и перестойных еловых насаждений приходится 36.9 и 51.2% всего запаса соответственно, тогда как на те же группы возраста сосновых лесов приходится только 12.9 и 30.8%. Это сравнение подчеркивает важность исследований способности именно еловых биогеоценозов Мурманской области к поглощению и аккумуляции атмосферного углерода.
Древостой является одним из ключевых углеродных пулов в лесных экосистемах мира, который хранит 367–394 Гт С (Mo et al., 2023). Оценка фитомассы древостоя, являющейся непосредственным резервуаром углерода, является важным направлением исследований (Wirth et al., 2004; Bonan, 2008; Thurner et al., 2014; Kenina et al., 2018; Nunes et al., 2020).
Считается, что существующие оценки запасов и поглощающей способности лесных экосистем нуждаются в повышении точности (Кутявин, 2018; Schepaschenko et al., 2021). Это возможно при получении дополнительных экспериментальных данных и длительном наземном мониторинге. Фитомасса лесного насаждения может изучаться путем рубки модельных деревьев, а затем статистической обработки значений массы различных фракций этих деревьев и их морфометрических характеристик и дальнейшей экстраполяции на всё насаждение. Фитомасса фракций деревьев поддаётся адекватной интерпретации путем применения различных математических моделей. Общепринятым подходом к расчету зависимостей фитомассы от дендрометрических характеристик деревьев, взятых по всем ступеням толщины в древостое, является регрессионная модель (Усольцев, 2016; Zianis et al., 2005). В дальнейшем построенные уравнения регрессии могут использоваться для расчетных оценок запасов фитомассы древостоя в сходных насаждениях (Трофимова и др., 2012; Осипов и др., 2019; Пеккоев и др., 2024; Шешницан и др., 2024; Kenina et al., 2018; Strîmbu et al., 2023; Merilä et al., 2024; Davidson et al., 2024; и др.).
Рельеф Мурманской области отличается своеобразием, накладывающим отпечаток на процессы динамики органического вещества в биогеоценозах. В регионе распространены морены, которые обусловливают холмистый, пересеченный рельеф местности. В зависимости от положения в рельефе складываются различные условия для развития лесных фитоценозов, что приводит к неоднородности продукционных показателей. Это обосновывает необходимость исследований фитомассы древостоя с учетом положения насаждений в пределах геохимически сопряженного ландшафта (Манаков, Никонов, 1981; Лукина и др., 2021).
Цель данного исследования – выявление возможности использования существующих моделей для оценки запасов фитомассы древостоев северотаежных ельников Мурманской области, сформированных в разных позициях ландшафта.
Накопленные в регионе и в схожих природных условиях экспериментальные данные по продуктивности северотаежных лесов являются основой для построения регрессионных моделей. Эти модели позволяют упростить оценку фитомассы, используя только основные дендрометрические характеристики – диаметр и высоту деревьев.
МАТЕРИАЛЫ И МЕТОДЫ
В пределах Мурманской области в соответствии с геоботаническим районированием выделяют таёжную зону, которая представлена в регионе лесотундрой и северотаёжной подзоной Кольско-Карельской подпровинции Североевропейской таёжной провинции (Геоботаническое районирование…, 1989). На формирование и динамику выделяемых зон влияет ряд особенностей природных условий. Ключевыми факторами, обусловливающими функционирование фоновых биогеоценозов региона, являются климат, почвообразующие породы и рельеф местности.
Район исследований расположен в Атлантико-Арктической зоне умеренного климата, отличающейся преобладанием теплых воздушных потоков, приходящих из северной Атлантики, а также холодных – из атлантического сектора Арктики (Алисов, 1956; Атлас…, 1971). Согласно классификации Кёппена-Гейгера, климат здесь субарктический (Dfc), характеризуется холодным летом и отсутствием сухих сезонов (Beck et al., 2023).
По лесорастительному районированию на территории Мурманской области выделены две лесорастительных зоны: таёжная зона, которая представлена северо-таёжным районом европейской части Российской Федерации, и зона притундровых лесов и редкостойной тайги – районом притундровых лесов и редкостойной тайги Европейско-Уральской части Российской Федерации. Общая площадь лесов, входящих в северо-таёжный район европейской части Российской Федерации, составляет 8819.9 тыс. га (Лесной план…, 2019).
Площадь земель, на которой расположены все леса в Мурманской области, составляет 10029.2 тыс. га, из них на землях лесного фонда – 9455.1 тыс. га. Покрытые лесной растительностью земли занимают 5144.4 тыс. га (Лесной план…, 2019).
В Мурманской области еловые леса, сложенные главным образом Picea obovata Ledeb. с единичной примесью P. abies (L.) Karst. в южных районах и несколько более значительным участием гибридогенной P. fennica (Regel.) Kom. в юго-западной части, занимают примерно 30% покрытой лесом площади (Малькова, Пешев, 1997; Лесной план…, 2019). Еловые фитоценозы характеризуются значительным участием березы (Betula pubescens s.l.). Экологическая амплитуда ели достаточно высока (Никонов, Лебедева, 1976; Цветков, Чертовский, 1979; и др.), что обусловливает разнообразие ассоциаций ельников.
Объекты исследования: ельники кустарничково-зеленомошные (в автоморфных и транзитных позициях ландшафта) и кустарничково-зеленомошные разнотравные (в аккумулятивных условиях) на юго-западе Мурманской области, сформированные на Al-Fe-гумусовых подзолах (Carbic Podzols), характеризующимися низким уровнем плодородия и pH. Древостой представлен елью финской (Picea fennica (Regel.) Kom), берёзой пушистой (Betula pubescens s.l.), сосной обыкновенной (Pinus sylvestris L.), реже осиной (Populus tremula L.). В подлеске встречается можжевельник (Juniperus sibirica Burgsd.), ива козья (Salix caprea L.), реже рябина Городкова (Sorbus gorodkovii Pojark.).
В травяно-кустарничковом ярусе доминирует черника (Vaccinium myrtillus L.), преобладают также вороника (Empetrum hermaphroditum Hagerup.), брусника (Vaccinium vitis—idaea L.). На всех исследованных пробных площадях (ПП) отмечен луговик (Avenella flexuosa (L.) Drejer), а также в меньшей степени встречалась ожика волосистая (Luzula pilosa (L.) Willd.), седмичник европейский (Trientalis europaea L.), марьянник лесной (Melampyrum sylvaticum L.), иван-чай узколистный (Chamaenerion angustifolium (L.) Scop.), золотарник обыкновенный (Solidago virgaurea L.). Мохово-лишайниковый ярус представлен в первую очередь зелеными мхами Pleurozium schreberi (Willd. ex Brid.) Mitt., Hylocomium splendens (Hedw.) Bruch et al., Dicranum sp., Ptilium crista-castrensis (Hedw.) De Not. В аккумулятивных позициях ландшафта отмечены виды, присущие более увлажненным местообитаниям: костяника (Rubus saxatilis L.) и ортилия однобокая (Orthilia secunda (L.) House), герань лесная (Geranium sylvaticum L.), голокучник обыкновенный (Gymnocarpium dryopteris (L.) Newman), лабазник вязолистный (Filipendula ulmaria (L.) Maxim.), перловник поникающий (Melica nutans L.).
Исследования выполнены в 2023-2024 гг. согласно методике, разработанной консорциумом «РИТМ Углерода» в рамках ВИП ГЗ «Единая национальная система мониторинга климатически активных веществ» (Методика …, 2023), на 15 ПП, расположенных на моренной гряде, вытянутой в меридиональном направлении, в автоморфных (5 ПП), транзитных (5 ПП) и аккумулятивных (5 ПП) позициях ландшафта. Транзитные ПП расположены на склоне южной экспозиции. Площадь каждой ПП – 0.25 га. В границах ПП проведен сплошной перечёт деревьев с измерением диаметра на высоте 1.3 м. В учёт включались все деревья более 6 см в диаметре. Высота оценивалась у не менее чем 15 экземпляров каждой породы. Далее, с помощью функций зависимости между диаметром и высотой деревьев, восстанавливали высоты всех деревьев. Возраст деревьев определяли с помощью подсчёта колец кернов, отобранных возрастным буравом Пресслера на высоте 20-30 см от шейки корня. Запасы древесины определялись по лесотаксационному справочнику (Лесотаксационный…, 1984).
Известно, что возраст и комплекс экологических и лесорастительных условий влияют на распределение фитомассы фракций каждого дерева (Нагимов и др., 2021). Также отмечают, что связывать массу структурных частей кроны с диаметром и высотой деревьев корректно только в древостоях сходного типа леса при незначительной возрастной гетерогенности во избежание существенных ошибок. Однако обилие работ по оценке фитомассы в разновозрастных древостоях, где в качестве метода описания фитомассы структурных частей деревьев применялась однофакторная регрессия с применением квадрата диаметра или произведения квадрата диаметра и высоты в качестве предиктора, демонстрирует применимость такого метода даже для древостоев со значительной возрастной гетерогенностью (Бобкова и др., 2010; Кузнецов, 2010; Осипов и др., 2022; и др.).
Для оценки запасов фитомассы деревьев на ПП были выбраны 4 модели.
- I. Региональная модель. Расчеты запасов фракций надземной фитомассы деревьев ели, сосны и берёзы проводились по коэффициентам аллометрического уравнения регрессии, основанных на региональных экспериментальных данных на примере коренных северотаёжных ельников кустарничково-зеленомошных и сосняков кустарничково-лишайниковых Мурманской области, произрастающих в автоморфных позициях ландшафта на Al-Fe-гумусовых подзолах (Лукина, Никонов, 1996). В качестве основного предиктора использовано произведение квадрата диаметра и высоты дерева, количество модельных деревьев каждой породы – 9 экземпляров.
В монографии представлены коэффициенты регрессионных уравнений для расчета запасов надземной фитомассы деревьев. Приведены коэффициенты регрессионного уравнения для расчёта следующих фракций фитомассы: хвоя (листва), ветви мелкие, ветви крупные, древесина ствола, кора ствола. Отсутствуют коэффициенты для расчёта подземной части деревьев. Оценка подземной части в данной статье дана как отношение подземной части фитомассы к надземной, согласно приведенным количественным значениям в монографии.
- II. Модели оценки запасов фитомассы ели, сосны и берёзы, разработанные в Финляндии, граничащей на западе с Мурманской областью (Repola et al., 2007). Основными предикторами являются диаметр и высота дерева. Учитываются следующие фракции фитомассы: хвоя (листва), ветви живые, корни крупные (>1 см), древесина ствола, кора, комлевая часть ствола (stump). Деревья отбирались в 36 еловых, 41 сосновых, 24 березовых насаждениях в широтном градиенте Финляндии, произрастающих в основном на минеральных почвах. Пробные площади закладывались в репрезентативных частях древостоев, как в молодых, так и спелых, также часть ПП включала экспериментальные участки по прореживанию и удобрению. Отбор модельных деревьев ели проводился в ельниках черничных (свежих на относительно плодородной почве) и ельниках кислично-черничных (относительно высокоплодородные), сосны – на бедных и плодородных почвах в сосняках вересковых, брусничных и черничных. Возраст ели варьировал от 18 до 161 года, березы от 11 до 97 лет, сосны от 13 до 145 лет. На каждой ПП отобрано по 4-5 модельных дерева. Уравнения линейные, имеют мультипликативную форму, для каждой отдельной фракции предусмотрены различные независимые переменные. Преимущество данной модели состоит в возможности расчёта подземной части деревьев. Авторами также отмечается, что модели обеспечивают надёжное прогнозирование биомассы компонентов дерева в широком диапазоне условий произрастания и насаждений в Финляндии (кроме листвы берёзы и подземной части деревьев). Применимость модели обусловлена сходными природно-климатическими условиями.
III. Двухфакторные регрессионные модели, основанные на материале по обобщению фактических данных фитомассы основных лесообразующих пород Евразии, охватывающих разные экорегионы и территории от Южной и Северной Европы, Урала до Средней и Юго-Восточной Азии (Усольцев, 2016). Учитываются следующие фракции фитомассы: хвоя (листва), ветви живые, корни, древесина ствола с корой. В основу моделей вошли многочисленные экспериментальные данные, в том числе по Кольскому полуострову (Чепурко, 1971, 1972; Манаков, Никонов, 1979, 1981; Алексеев, Рак, 1985; Лукина, Никонов, 1991, 1996). Целью работы (Усольцев, 2016) было создание унифицированных регрессионных моделей фитомассы в масштабе континента по ряду древесных пород с учетом зональности и континентальности. Оценки фитомассы деревьев для наземной таксации проводятся на основе диаметра и высоты. В работе даны регрессионные модели для ели, сосны и березы с учётом данных по Мурманской области.
- IV. Модели расчета пула запаса углерода в биомассе деревьев по регрессионному уравнению с традиционными предикторами – диаметром и высотой дерева, приведенные в Приложении к методике количественного определения объема поглощений парниковых газов, утвержденной приказом Минприроды России от 27.05.2022 №371 (Приказ…, 2022). Определялись надземная, подземная и общая фитомасса. Необходимо отметить, что в документе имеются таблицы для расчёта запасов углерода в фитомассе деревьев через запас древесины, однако в данном исследовании рассматриваются только модели регрессионных уравнений с дендрометрическими характеристиками в качестве предикторов.
Краткая характеристика и различия моделей приведены в табл. 1.
Таблица 1. Краткая характеристика моделей
| Модель | Авторы, год | Рассчитываемые части фитомассы | ||
| ель | сосна | берёза | ||
| I | Н. В. Лукина,
В. В. Никонов (1996) |
надземная по фракциям | надземная по фракциям | надземная по фракциям |
| II | J. Repola et al. (2007) | общая по фракциям | общая по фракциям | общая по фракциям |
| III | В. А. Усольцев (2016) | общая по фракциям | общая по фракциям | общая по фракциям |
| IV | Приказ Минприроды… (2022) | общая | надземная | общая |
Примечание: общая фитомасса подразумевает возможность расчёта подземной части фитомассы наряду с надземной.
В качестве эталонной для расчёта запасов фитомассы в автоморфных позициях ландшафта выбрана региональная модель I, так как данные получены непосредственно в еловых биогеоценозах Мурманской области, сформированных на возвышенностях. Фитомасса коры и фитомасса стволовой древесины суммировались и приводятся как одна фракция. Для характеристики изменчивости оценок фитомассы разными моделями применялся коэффициент вариации (CV), выраженный в процентах (%). В качестве метода сравнения результатов для сопоставимых оценок выбран парный двухвыборочный t-тест для средних.
Для выполнения поставленной цели выбраны ельники кустарничково-зеленомошные и кустарничково-зеленомошные разнотравные.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Запасы фитомассы древостоя. Для оценки фитомассы древостоя в ненарушенных условиях были выбраны ельники различных (IV–X) классов возраста, Vа класса бонитета. Таксационная характеристика древостоев на пробных площадях представлена в табл. 2. Максимальные значения фитомассы отмечаются в автоморфных и аккумулятивных позициях ландшафта, снижаясь к транзитным условиям.
Таблица 2. Таксационная характеристика древостоев на пробных площадях
| ПП | Порода | Средние | Возраст ели, лет | Густота живых деревьев, шт. га-1 | Сумма площадей сечения, м2 га-1 | Запас древесины, м3 га-1 | Состав древостоя | |
| d, см | h, м | |||||||
| Автоморфные условия | ||||||||
| 30-23 | Е | 18.3 | 11.0 | 50-300 | 740 | 12.2 | 76 | 8Е2Б |
| Б | 10.3 | 7.6 | 4.1 | 19 | ||||
| 31-23 | Е | 16.5 | 10.0 | 60-240 | 604 | 8.1 | 51 | 7Е3Б |
| Б | 11.2 | 8.0 | 3.6 | 20 | ||||
| 32-23 | Е | 16.1 | 9.9 | 55-265 | 916 | 7.2 | 45 | 6Е4Б |
| Б | 10.2 | 7.5 | 6.8 | 34 | ||||
| 33-23 | Е | 19.4 | 11.4 | 145-255 | 856 | 9.1 | 59 | 6Е2С2Б |
| Б | 10.5 | 7.6 | 4.4 | 22 | ||||
| С | 14.3 | 10.3 | 3.5 | 21 | ||||
| 34-23 | Е | 16.1 | 9.9 | 55-155 | 740 | 6.5 | 40 | 5Е1С3Б1Ос |
| Б | 10.5 | 7.7 | 4.5 | 20 | ||||
| С | 18.0 | 11.9 | 1.1 | 7 | ||||
| Ос | 12.8 | 8.7 | 1.2 | 7 | ||||
| Транзитные условия | ||||||||
| 35-23 | Е | 18.4 | 11.1 | 70-275 | 840 | 6.3 | 48 | 6Е4Б |
| Б | 11.2 | 8.3 | 7.6 | 28 | ||||
| 36-23 | Е | 18.1 | 11.1 | 80-310 | 844 | 8.5 | 53 | 7Е3Б |
| Б | 10.0 | 7.8 | 6.1 | 26 | ||||
| 37-23 | Е | 14.8 | 9.3 | 55-170 | 816 | 3.6 | 22 | 4Е1С5Б |
| Б | 10.0 | 7.8 | 6.0 | 29 | ||||
| С | 17.0 | 11.0 | 1.3 | 9 | ||||
| 38-23 | Е | 15.8 | 9.8 | 80-210 | 840 | 5.5 | 33 | 5Е5Б |
| Б | 11.0 | 8.2 | 6.4 | 32 | ||||
| 39-23 | Е | 15.6 | 9.8 | 80-210 | 896 | 4.7 | 37 | 5Е1С4Б |
| Б | 11.1 | 8.3 | 8.3 | 28 | ||||
| С | 18.4 | 11.0 | 1.5 | 10 | ||||
| Аккумулятивные условия | ||||||||
| 40-23 | Е | 14.9 | 9.2 | 70-230 | 1512 | 7.3 | 45 | 5Е5Б+С |
| Б | 10.2 | 8.8 | 11.1 | 53 | ||||
| С | 13.3 | 9.8 | 0.1 | 1 | ||||
| 41-23 | Е | 15.9 | 9.7 | 50-95 | 1404 | 4.6 | 28 | 4Е6Б |
| Б | 8.9 | 8.2 | 8.0 | 36 | ||||
| 42-23 | Е | 12.8 | 8.4 | 90-110 | 1452 | 13.2 | 69 | 8Е2Б |
| Б | 8.8 | 8.2 | 4.3 | 18 | ||||
| 43-23 | Е | 12.2 | 8.2 | 65-100 | 1076 | 7.8 | 41 | 7Е1С2Б |
| Б | 8.5 | 8.0 | 3.5 | 12 | ||||
| С | 21.0 | 12.0 | 0.7 | 5 | ||||
| 44-23 | Е | 13.9 | 8.9 | 80-100 | 1076 | 7.9 | 45 | 6Е2С2Б |
| Б | 8.8 | 8.1 | 4.1 | 16 | ||||
| С | 16.5 | 10.8 | 3.1 | 10 | ||||
Примечания: d — средний диаметр на высоте 1.3 м, h — средняя высота; формула состава древостоя дана по запасам.
Автоморфный ландшафт. Фитомасса древостоя на исследованных пробных площадях, приуроченных к автоморфным позициям ландшафта, в среднем по всем моделям (табл. 1) варьирует от 66.1 ± 5.1 (модель IV, CV = 13.4%) до 70 ± 6.3 т га-1 (модель III, CV = 20.3%) (рис. 1). При этом отмечаются одинаковые значения коэффициента вариации при применении моделей I и II (CV = 18.4%, при 68.1 ± 5.6 и 66.8 ± 5.5 т га-1, соответственно). Значения фитомассы древостоев в среднем по всем моделям изменяются от 54 ± 0.3 до 82.2 ± 1.6 т га-1.
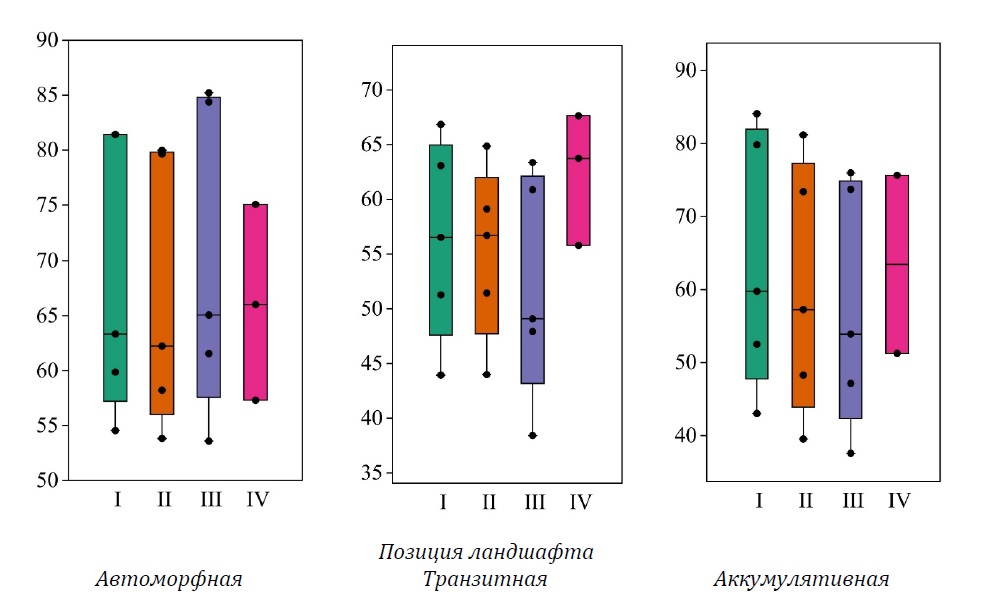
Рисунок 1. Фитомасса древостоя в ельниках (т га-1), сформированных в различных позициях ландшафта, рассчитанная по нескольким моделям: I, II, III, IV (см. табл. 1)
Обнаружено, что фитомасса древостоя, рассчитанная по модели II, всегда меньше (различия статистически значимы, p = 0.002, n = 5), чем по региональной модели (I), которую для автоморфных позиций ландшафта мы приняли за «эталонную», на 1.3–2.7%. Различия в фитомассе, оцененной по региональной модели и моделям III и IV, демонстрируют более выраженную изменчивость и разнонаправленность, в отличие от оценок по модели II. Так, III модель дает несколько меньшие значения для ПП 34-23 и 32-23 (меньше на 1.8, 2.8% соответственно) и несколько большие для ПП 30-23, 33-23 и 31-23 (больше на 3.6, 4.7 и 8.7% соответственно), однако различия статистически незначимы (p = 0.25, n = 5). Запас фитомассы древостоя по модели IV определялся только для тех площадок, где отсутствовала сосна, поскольку в приказе Минприроды (Приказ…, 2022) не содержится коэффициентов аллометрического уравнения для расчета подземной фитомассы сосны. Оценки по данной модели были на 4.3 и 7.8% ниже (различия также статистически незначимы, p = 0.51, n = 3) региональных оценок на ПП 30-23 и 31-23, и на 4.2% больше на ПП 32-23.
В автоморфных условиях на основную лесообразующую породу – ель – приходится в среднем по всем моделям на каждой ПП от 34.1±2 до 65.4±4 т га-1 биомассы. Вклад берёзы в запасы фитомассы ельников значителен и варьирует в среднем по всем моделям от 14.4±2 до 24.3 ± 3.5 т га-1. На двух ПП (33-23 и 34-23) отмечены деревья сосны, фитомасса которых варьирует от 4.7 ± 0.1 до 13.9 ± 0.1 т га-1.
Фитомасса еловой части древостоя (рис. 2) варьирует от 39.4 ± 4.6 (IV, CV = 26%) до 54.7 ± 6.2 т га-1 (III, CV = 25.3%), несколько более высокий коэффициент вариации отмечен для региональной модели (CV = 25.6%, при 48.3 ± 5.5 т га-1) и для модели II (CV = 26.3%, при 45.9 ± 5.4 т га-1).
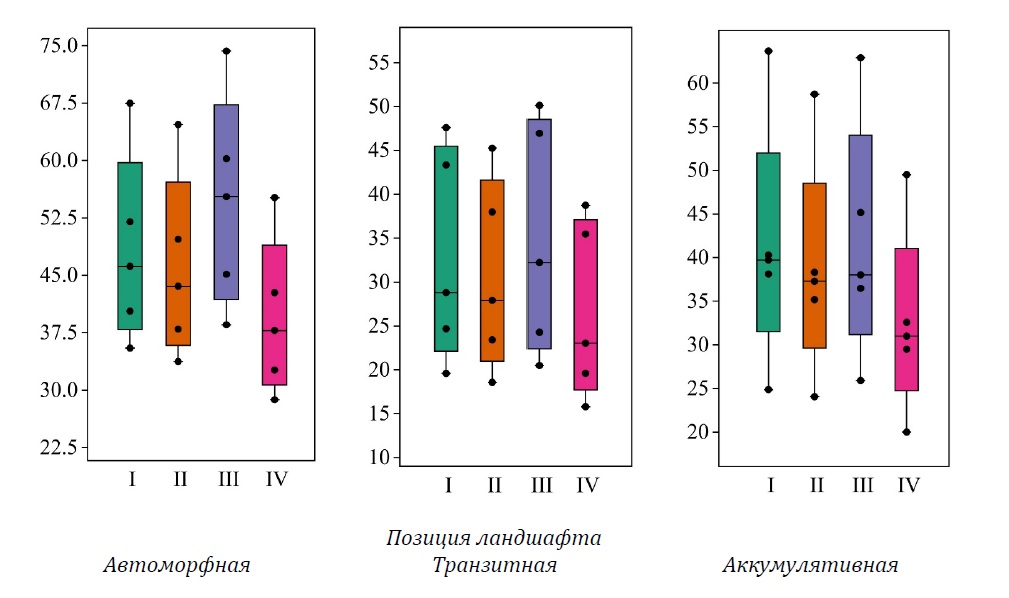
Рисунок 2. Фитомасса еловой части древостоя (т га-1), сформированной в различных позициях ландшафта, рассчитанная по нескольким моделям: I, II, III, IV (см. табл. 1)
Отмечено занижение оценок относительно региональных значений фитомассы деревьев ели по модели II на 4.1–5.8%. По модели III, наоборот, наблюдается завышение на 8.5–19.8%. На всех ПП по модели IV отмечается занижение на 17.8–19% (различия статистически значимы по всем моделям, p = 0.002, 0.014, 0.0005 для II, III, IV соответственно при n = 5).
Общая фитомасса березовой части древостоя (рис. 3) варьирует от 11.5 ± 1.2 т га-1 (III, CV = 24%) до 23.3 ± 2.6 (IV, CV = 24.9%). Региональная (I) и финская (II) модели дают близкие значения (I – 16.2 ± 1.8 т га-1, II – 17.1 ± 1.8 т га-1), значения фитомассы, рассчитанные по модели II, демонстрируют самый низкий коэффициент вариации (I – CV = 24.2%, II – CV = 23.7%).
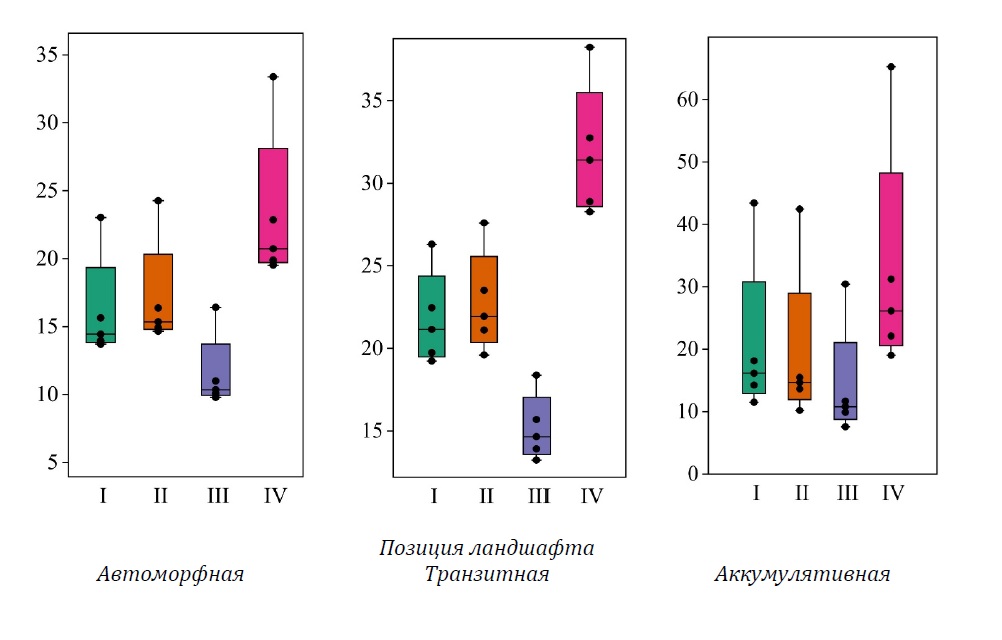
Рисунок 3. Фитомасса берёзовой части древостоя (т га-1), сформированного в различных позициях ландшафта, рассчитанная по нескольким моделям: I, II, III, IV (см. табл. 1)
Варьирование оценок фитомассы деревьев берёзы, рассчитанных по разным моделям, более значительно по сравнению с оценками для елового компонента древостоя. Такие значения связаны с вероятной ошибкой, допущенной в приказе Минприроды (2022). Результаты расчётов только надземной фитомассы по модели IV систематически до 56.1% превышают значения, полученные по региональной модели. В связи с этим превышения по общей фитомассе берёзы, рассчитанные как сумма надземной и подземной частей дерева, достигают 46.1% по сравнению с региональной моделью. Существенные занижения общей фитомассы берёзы отмечены для модели III (на 27.7–29.7%), это может быть связано с тем, что экспериментальные данные из Мурманской области по данной породе не были включены в модель. Оценки, полученные по модели II, наиболее близки к региональным: завышения на 4.7–7% относительно региональных значений (различия статистически значимы по всем моделям, p = 0.001, 0.001, 0.002 для II, III, IV соответственно при n = 5).
Значения общей фитомассы сосновой части древостоя, полученные по трем моделям (I–III), варьируют незначительно. Фитомасса сосны по модели II превышает оценки по региональной модели на 1–3.1%, а по модели III – на 1.6–2.1%.
Согласно результатам сравнения оценок запасов фитомассы на каждой ПП по моделям II, III и IV с «эталонными» региональными оценками запасов фитомассы (модель I) с помощью парного t-теста для средних, общая фитомасса древостоя ели, полученная с помощью модели III, достоверно сопоставима на ПП 32-23 и 34-23 (p = 0.1, 0.07 соответственно). Общая фитомасса древостоя сосны, полученная с помощью модели II, сопоставима с запасами по региональной модели на ПП 33-23 (p = 0.6).
Таким образом, для оценок общей фитомассы компонентов древостоев основных видов деревьев в северотаёжных ельниках Мурманской области, сформированных в автоморфных позициях ландшафта, наиболее подходящей моделью является финская модель (II), что подтверждает самая незначительная разница как по общим запасам фитомассы древостоя, так и отдельно по видам древесных растений. Модель построена на значительном экспериментальном материале, отобранном как в Северной Финляндии, так и в Южной, в различных лесорастительных условиях. Это, вместе с результатами сравнения оценок с региональной моделью, указывает на широкую применимость модели для северотаёжных ельников.
Модель III (Усольцев, 2016), вероятно, можно использовать для оценок фитомассы еловой части древостоя, что подтверждается статистическим анализом. Однако, учитывая значительные отличия в запасах фитомассы елей, а также систематические занижения оценок фитомассы берёзовой части древостоя, которая является основной сопутствующей породой в ельниках Кольского полуострова, можно сделать вывод о невозможности использования данной модели для оценок запасов фитомассы смешанных древостоев в еловых биогеоценозах региона. Также пример таёжных еловых лесов, формирующихся на северном пределе распространения деревьев ели, указывает на ограничения использования подобных «всеобщих» моделей и необходимость дальнейших исследований, для включения новых экспериментальных данных в подобные модели, т.к. различия климатических, почвенных и иных условий могут повлиять на качество полученных оценок.
Использование модели IV не обосновано по причине значительных отклонений, отмеченных при сравнении оценок, полученных по региональной и другим моделям.
Транзитный и аккумулятивный ландшафты. Поскольку не существует региональных моделей для оценок запасов фитомассы древостоев, сформированных в транзитных и аккумулятивных позициях ландшафта, для достижения поставленной цели мы можем рассматривать используемые нами модели в зависимости от того, какие тенденции выявляются при использовании существующих моделей друг относительно друга и сопоставлять с тенденциями, отмеченными в автоморфных условиях.
В транзитных позициях ландшафта запасы фитомассы (рис. 1) в среднем при использовании моделей варьируют от 51.9 ± 4.6 (III, CV = 19.7%) до 62.4 ± 3.5 т га-1 (IV, CV = 9.7%), модели I и II демонстрируют средние значения коэффициента вариации и близкие средние запасы (CV = 16.2 и 14.3 %, при 56.3 ± 4.1 и 55.2 ± 4.5 т га-1 соответственно). Запасы изменяются от 42.1 ± 1.8 т га-1 (ПП 37-23) до 65.7 ± 1 т га-1 (ПП 36-23) в среднем на ПП при применении всех моделей.
В транзитных условиях в среднем по всем моделям на пяти ПП фитомасса еловой части древостоя варьирует от 23 ± 1.2 до 45.4 ± 2.4 т га-1. Фитомасса берез варьирует в среднем по всем моделям от 20.2 ± 3.2 до 27.6 ± 4.1 т га-1. На двух ПП (37-23 и 39-23) присутствует сосна, фитомасса которой в среднем по трём моделям варьирует от 3.3 ± 0.1 до 5.9 ± 0.3 т га-1.
Фитомасса елей в транзитных условиях (рис. 2) варьирует от 26.5 ± 4.5 (модель IV, CV = 37.9%) до 34.8 ± 5.9 т га-1 (III, CV = 38.1%), сходный коэффициент вариации отмечен для региональной модели (CV = 36.9%, при 32.8 ± 5.4 т га-1) и для модели II (CV = 35.5%, при 30.6 ± 4.9 т га-1). Запасы фитомассы березовой части (рис. 3) варьируют от минимальных 15.2 ± 0.9 т га-1 (III, CV=13.2%) до максимальных 31.9 ± 1.8 т га-1 (IV, CV=12.5%). При использовании моделей I и II снова получены близкие значения (I – 21.8 ± 1.3 т га-1, II – 22.8 ± 1.4 т га-1) и сходные коэффициенты вариации (I – CV=13%, II – CV=13.4%).
В исследованных ельниках, сформированных в аккумулятивных позициях ландшафта, фитомасса древостоя (рис. 1) варьирует от 57.7 ± 7.5 (модель III, CV = 29%) до 63.8 ± 7.9 т га-1 (модель I, CV = 27.6%). Наибольший коэффициент вариации отмечается у модели III. По IV модели CV = 27.2% при 63.4 ± 12.2 т га-1 фитомассы. Модель II демонстрирует умеренные результаты (59.9 ± 7.7 т га-1, CV = 28.8%).
В среднем по всем моделям на ПП в аккумулятивных условиях запасы фитомассы ели варьируют от 23.7 ± 1.3 до 58.7 ± 3.3 т га-1. Запасы фитомассы берёзы варьируют в среднем по всем моделям от 12.1 ± 2.5 до 45.4 ± 7.2 т га-1. На трех ПП (40-23, 43-23 и 44-23) присутствует сосна, фитомасса которой в среднем по трём моделям варьирует от 0.4 ± 0.01 до 6 ± 0.2 т га-1.
Фитомасса еловой части древостоя в биогеоценозах, сформированных в аккумулятивных позициях ландшафта (рис. 2), варьирует от 32.5 ± 4.8 (модель IV, CV = 32.8%) до 41.7 ± 6.1 т га-1 фитомассы (III, CV = 32.9%). Сходный коэффициент вариации отмечен для модели II (CV=32.4%, при 38.7 ± 5.6 т га-1). Фитомасса елей, рассчитанная по модели I, составила 41.3 ± 6.3 т га-1 (CV=33.9%). Запас фитомассы берёз (рис. 3) варьирует от 14.1 ± 4.1 (модель III, CV=65.9%) до 32.8 ± 8.4 т га-1 (модель IV, CV=57.2%). При использовании моделей I и II получены близкие значения фитомассы (модель I – 20.7 ± 5.8 т/га, II – 19.3 ± 5.9 т га-1), но при использовании модели II коэффициент вариации ближе к значению, полученному для модели III (модель I – CV=62.5%, II – CV=67.9%).
В транзитных и аккумулятивных ландшафтах, как и в ельниках, сформированных в автоморфных условиях, сохраняются общие тенденции, обусловленные разницей в оценках по модели III относительно региональной. Сохраняется тенденция к систематическому и значительному занижению запасов фитомассы берёзовой части древостоя, рассчитанных по модели III, по сравнению с оценками по моделям I и II (соответственно, на 30.3 и 33.3% в среднем в транзитных условиях и на 32.8 и 26.6% – в аккумулятивных).
Выраженные тенденции в различиях по фитомассе отмечаются и для еловой части древостоя. Среднее превышение запасов фитомассы, рассчитанных по модели III, относительно расчетов по модели I варьирует от 13.2% в автоморфных условиях до 5.7 и 6.3% в транзитных и аккумулятивных позициях ландшафта, а относительно расчётов по модели II изменяется от 19.2% в автоморфной позиции до 12.8 и 14% в транзитных и аккумулятивных.
Поскольку сохраняется значительное превышение оценок фитомассы березовой части древостоя по модели IV относительно других моделей – до 2 раз в транзитных и аккумулятивных элементарных ландшафтах – данная модель не может быть использована для оценки запасов фитомассы ельников северной тайги.
Полученные значения коэффициентов вариации для ПП в автоморфных позициях близки к полученным значениям CV в транзитных условиях (13–20.3% в автоморфных и 10–19.7% в транзитных), что может указывать на естественную однородность запасов фитомассы коренных разновозрастных ельников вне зависимости от положения в рельефе.
Более высокий CV в аккумулятивных условиях (27–29%) может свидетельствовать о структурной гетерогенности ельников различного сукцессионного статуса: отмечено два одновозрастных (V-VI класса возраста) и три разновозрастных древостоя (IV, VII класса возраста). При этом можно отметить крайне неравномерный вклад берёзы в аккумулятивных позициях (CV более 60–67%). Это также должно свидетельствовать о сукцессионной динамике леса на исследованных ПП, значительный вклад в которую вносят деревья этой породы.
Фракции фитомассы деревьев ельников в автоморфных позициях ландшафта
Более детальный анализ структуры фитомассы позволяет определить, какая фракция древостоев аккумулирует больше органического вещества и, следовательно, углерода. Полученные результаты указывают на неоднозначность в закономерностях продуцирования органического вещества. Согласно региональной модели, в структуре фитомассы прослеживается следующий ряд по участию в запасах: стволовая древесина (с корой) > корни > ветви > хвоя (листва). При общем сохранении такого ряда участия в запасах фитомассы доли фракций различаются по всем рассмотренным моделям (рис. 4).
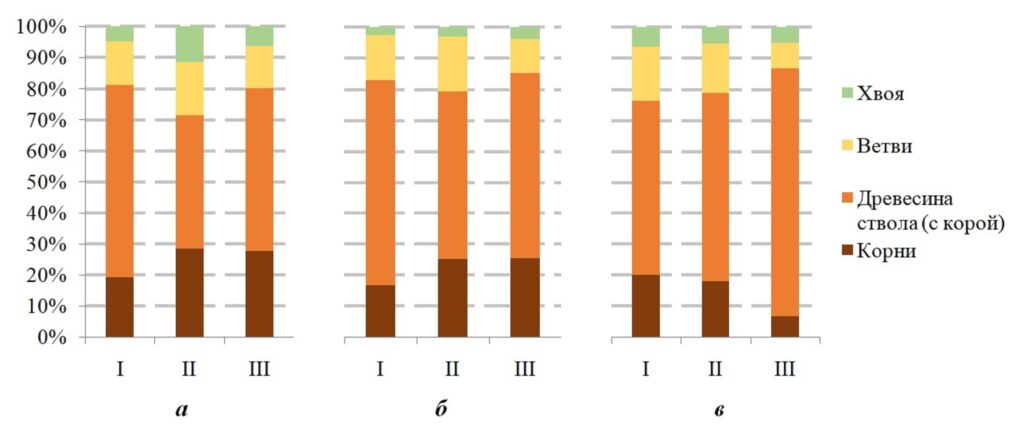
Рисунок 4. Доля фракций в запасе фитомассы (а) елового, (б) берёзового и (в) соснового компонента древостоя, рассчитанная по моделям I, II, III (см. табл. 1)
Так, если по модели I на стволовую древесину ели в коре приходится около 62% всего запаса, то по модели II – 42–45%, а по модели III – 52–54%. При этом результаты, полученные по региональной модели, подтверждаются исследованиями в еловых биогеоценозах Мурманской области (Манаков, Никонов, 1981; Лукина, Никонов, 1996). Так, на запасы фитомассы древесины в ельниках, сформированных в автоморфных позициях ландшафта, приходится от 59 до 64%. Самая высокая доля в запасах фитомассы стволовой древесины берёзы получена по региональной модели I (66.5 ± 0.1%), тогда как по моделям II и III – 54.1± 1.1 и 59.9 ± 0.6% соответственно. Согласно литературным данным, доля фракций фитомассы берёзы в структуре запасов может варьировать. Так, доля древесины ствола в фитомассе берёзы северотаёжных ельников Мурманской области может варьировать от 57 до 76% (Манаков, Никонов, 1981; Лукина, Никонов, 1996).
На запасы фитомассы корней ели, рассчитанные по моделям II и III, приходится от 27 до 30%, тогда как по региональной модели значительно меньше – 19%. Согласно уже упомянутым работам, в исследуемом регионе на запасы фитомассы корней ели в ельниках Мурманской области приходится от 19 до 23% всей фитомассы ели, как в автоморфных, так и в транзитных условиях (Манаков, Никонов, 1981). Подобные результаты отмечались в ельниках средней тайги Финляндии: 23–24%, тогда как в северотаежных лесах доля корней в фитомассе ели доходила до 24.6% (Merilä et al., 2024). В Мурманской области столь высокая (от 34 до почти 40 %) доля корней в фитомассе ели отмечалась только в аккумулятивных условиях, но в ельниках, на торфяно-болотных почвах (Манаков, Никонов, 1981). Доля корней в среднетаёжных ельниках Карелии оказалась ниже. В зависимости от типа сообщества (рассматривалось 7 типов, представляющих эдафо-фитоценотический ряд), доля корней от общей фитомассы в ельниках средневозрастных варьировала от 17 до 19% (Казимиров, Морозова, 1973). В том же исследовании показано, что вне зависимости от возраста ельника черничного, сформированного на подзолистых иллювиально-гумусово-железистых почвах, отношение корней к общей фитомассе стабильно варьирует в подобных же пределах (18–19%). В среднетаежном коренном ельнике черничном в Республике Коми, произрастающем на подзолах, сформированных на покровных суглинках, доля корней ели составляет около 23% (Осипов и др., 2019). Стоит отметить, что авторы модели II упоминают ограниченную применимость их модели при оценках подземной части деревьев.
Запас фитомассы корней берёзы по моделям II и III превышает региональные значения в среднем почти на 9%.
При расчете фитомассы по региональной модели доля фитомассы корней бралась как неизменная величина для ели, берёзы и сосны: 19, 17 и 20%, соответственно). Однако если для ели это оправдано незначительной изменчивостью вклада данной фракции в фитомассу дерева, то при расчёте фитомассы берёзы это может привести к значительным ошибкам, поскольку согласно исследованиям (Манаков, Никонов, 1981) доля корней в фитомассе берёзы в ельниках Мурманской области может варьировать от 17 до 28%.
Результаты, полученные с помощью модели III для сосны, указывают на нарушение закономерности, выявленной для остальных пород, также и по другим моделям, в том числе по расчетам фитомассы сосны. Так, согласно модели III, доля ветвей в запасе фитомассы сосны сопоставима с долей корней. По данной модели отмечено превышение доли стволовой древесины в запасах сосны (80.1 ± 0.1%) в среднем на 24% относительно региональной модели (56.3 ± 0.2%), тогда как по модели II – 61 ± 0.1% – превышение по данной фракции незначительно. Доля корней в общей фитомассе сосны по модели III на 13% меньше, чем по региональной: 7 % и 20%, соответственно.
Неоднозначность в оценках доли корней деревьев свидетельствует о необходимости получения экспериментальных данных о вкладе корней в запас фитомассы ельников.
Выявлена статистически значимая сопоставимость оценок надземной фитомассы ели (сумма всех оцениваемых фракций без корней) по модели III с региональной моделью на ПП 30-23…34-23 (p = 0.3…0.9). Запасы по модели III (39.4 ± 4.5 т га-1) на 0.4 ± 0.6 т га-1 выше региональных (39 ± 4.5 т га-1). Также статистически достоверно сопоставимы с региональными оценками значения общей фитомассы стволовой древесины и коры ели, определенной по модели III для ельников на ПП 31-23…33-23 (p = 0.1…0.9).
Запасы надземной фитомассы, рассчитанные с применением модели II, составляли 32.4 ± 4.2 т га-1, что в среднем существенно ниже (на 6.6 ± 0.4 т/га) по сравнению с оценками по региональной модели. Статистически достоверно сопоставимы с региональными оценками значения надземных запасов фитомассы сосны на ПП 33-23 по модели II (p = 0.07), а также запасы хвои и ветвей сосны (p = 0.07 и p = 0.5 соответственно). Сопоставимые с региональными оценки обнаружены для фитомассы листвы и ветвей берёзы, полученные по модели II (p = 0.09 и 0.7 соответственно).
ЗАКЛЮЧЕНИЕ
Анализ сравнения полученных данных с литературными источниками позволяет сделать вывод о возможности применения модели II (разработанной в Финляндии) для оценки фитомассы древостоя в северотаёжных еловых лесах Мурманской области, поскольку данная модель обеспечивает наиболее точную и стабильную оценку общего запаса органического вещества во всех ландшафтных позициях. Оценки по данной модели демонстрируют наиболее предсказуемый разброс значений фитомассы древостоя во всех типах элементарного ландшафта для ельников кустарничково-зеленомошных, сформированных на подзолах Al-Fe-гумусовых, поскольку модель калибрована с учетом пространственной вариабельности в сходных природно-климатических условиях. Однако при оценке структуры фитомассы отмечаются значительные различия, что критично при исследованиях, учитывающих фракции фитомассы древостоя. Модель является наиболее сбалансированным, предсказуемым и практически применимым компромиссом для оценки углеродных запасов в северотаёжных ельниках Мурманской области на текущий момент.
Модель В. А. Усольцева (III) дает значительные и разнонаправленные расхождения в оценках фитомассы. Выявленные систематические занижения запасов фитомассы берёзы относительно региональной и «финской» моделей могут привести к недоучёту запасов ключевой сопутствующей породы в северотаёжных ельниках Мурманской области и, соответственно, при оценках общих запасов фитомассы на ПП. Для специфических условий региона данная модель ограниченно пригодна: при применении этой модели получены сопоставимые с региональными значениями оценки запасов фитомассы ели (надземной фитомассы, фитомассы стволовой древесины) в автоморфных условиях.
Модель IV (Приказ…, 2022) в её текущем виде обладает ограниченной научной и практической ценностью для достоверной оценки фитомассы древостоя в северотаёжных ельниках. Её применение приведёт к значительным систематическим отклонениям, особенно для берёзы и сосны. Для целей научно-обоснованного мониторинга углеродных пулов и калибровки дистанционных методов в данном регионе модель не рекомендована. Требуется её доработка с учётом региональной специфики.
Отсутствие надёжных аллометрических уравнений для корней ели, берёзы и сосны –существенный пробел, ограничивающий точность учета запасов фитомассы древостоя. Значительные расхождения в оценках вклада фракций фитомассы в общий запас, а также высокая изменчивость в оценках фитомассы берёзы указывают на необходимость дополнительных экспериментальных исследований с целью уточнения региональных аллометрических моделей для достоверной количественной оценки роли северотаежных ельников Мурманской области в углеродном балансе.
БЛАГОДАРНОСТИ
Авторы выражают искреннюю благодарность сотрудникам Лаборатории наземных экосистем ИППЭС КНЦ РАН за помощь и участие в сборе полевых материалов: Сухаревой Т. А., Ершову В. В., Ивановой Е. А., Живову Д. А., Сидоровой Е. А., особая благодарность д.б.н., члену-корреспонденту РАН Лукиной Н.В. за ценные советы и комментарии.
ФИНАНСИРОВАНИЕ
Исследование выполнено в рамках Государственного задания Института проблем промышленной экологии Севера ФИЦ КНЦ РАН (рег. номер 125021402277-1) «Структурно-функциональная организация и динамика наземных экосистем Евро-Арктического региона», а также в рамках реализации важнейшего инновационного проекта государственного значения (ВИП ГЗ) «Единая национальная система мониторинга климатически активных веществ».
СПИСОК ЛИТЕРАТУРЫ
Алексеев В. А., Рак Л. Д. Признаки ослабления елей под влиянием атмосферного загрязнения // Лесоведение. 1985. № 5. С. 37–43.
Алисов Б. П. Климат СССР. М.: МГУ, 1956. 125 с.
Атлас Мурманской области. М., 1971. 33 с.
Бобкова К. С., Кузнецов М. А., Манов А. В., Галенко Э. П., Тужилкина В. В. Фитомасса древостоев ельников чернично-сфагновых на болотно-подзолистых почвах Европейского Северо-Востока // Известия высших учебных заведений. Лесной журнал. 2010. № 1. С. 19–26.
Геоботаническое районирование Нечерноземья Европейской части РСФСР / Отв. ред. В. Д. Александрова. Л.: Наука, 1989. 61 с.
Зайцева И. В., Кобяков К. Н., Никонов В. В., Смирнов Д. Ю. Коренные старовозрастные леса Мурманской области // Лесоведение. 2002. № 2. С. 14–22.
Казимиров Н. И., Морозова Р. М. Биологический круговорот веществ в ельниках Карелии. Л.: Наука, 1973. 175 с.
Кузнецов М. А. Динамика содержания органического углерода в заболоченных ельниках средней тайги: Дисс. … канд. биол. наук (спец. 03.02.08). Сыктывкар, 2010. 141 с.
Кутявин И. Н. Сосновые леса Северного Приуралья: строение, рост, продуктивность / Отв. ред. К. С. Бобкова. Сыктывкар: ИБ Коми НЦ УрО РАН, 2018. 176 с. DOI: 10.31140/book-2018-02
Лесной план Мурманской области. В 2-х томах. Мурманск, 2019. 106 с.
Лесотаксационный справочник по Северо-Западу СССР. Ленинград: ЛТА, 1984. 320 с.
Лукина Н. В., Гераськина А. П., Кузнецова А. И., Смирнов В. Э., Горнов А. В., … & Басова Е. В. Функциональная классификация лесов: актуальность и подходы к разработке // Лесоведение. 2021. № 6. С. 566–580. DOI: 10.31857/S0024114821060085
Лукина Н. В., Никонов В. В. Изменение первичной продуктивности ельников под влиянием техногенного загрязнения на Кольском полуострове // Лесоведение. 1991. № 4. С. 37–45.
Лукина Н. В., Никонов В. В. Биогеохимические циклы в лесах Севера в условиях аэротехногенного загрязнения. В 2-х ч. Ч. 1. Апатиты: Изд-во Кольского научного центра РАН, 1996. 213 с.
Лукина Н. В., Смирнов В. Э., Тебенькова Д. Н., Данилова М. А., Тихонова Е. В., … & Ручинская Е. В. Роль старовозрастных лесов в аккумуляции и хранении углерода // Известия Российской академии наук. Серия географическая. 2023. Т. 87. № 4. C. 536–557. DOI: 10.31857/S2587556623040064
Малькова Т. Н., Пешев Н. Г. Лесные ресурсы Кольского Севера: эколого-экономические аспекты лесопользования. Апатиты: Изд-во КНЦ РАН, 1997. 84 с.
Манаков К. Н., Никонов В. В. Первичная биологическая продуктивность ельников Кольского полуострова // Ботанический журнал. 1979. Т. 64. № 2. С. 232–241.
Манаков К. Н., Никонов В. В. Биологический круговорот минеральных элементов и почвообразование в ельниках Крайнего Севера. Ленинград: Наука, 1981. 196 с.
Методика полевых работ по таксации леса на постоянных пробных площадях в рамках реализации инновационного проекта государственного значения «Углерод в экосистемах: мониторинг». М.: ЦЭПЛ РАН, 2023. 32 с.
Нагимов З. Я., Артемьева И. Н., Шевелина И. В., Нагимов В. З. Оценка рангового положения деревьев в древостое при исследовании их фитомассы // Успехи современного естествознания. 2021. № 7. С. 20–25. DOI: 10.17513/use.37657
Никонов В. В., Лебедева Р. М. Ель и еловые леса в центральной части Кольского полуострова // Изучение растительных ресурсов Мурманской области. Апатиты: Изд-во Кольского филиала АН СССР, 1976. С. 53–64.
Осипов А. Ф., Кутявин И. Н., Манов А. В., Кузнецов М. А., Бобкова К. С. Запасы и структура фитомассы древостоев северотаежных сосняков Республики Коми // Известия высших учебных заведений. Лесной журнал. 2022. № 4. С. 25–38.
Осипов А. Ф., Тужилкина В. В., Дымов А. А., Бобкова К. С. Запасы фитомассы и органического углерода среднетаежных ельников при восстановлении после сплошнолесосечной рубки // Известия Российской академии наук. Серия биологическая. 2019. № 2. С. 215–224. DOI: 10.1134/S0002332919020103
Пеккоев А. Н., Мошников С. А., Ромашкин И. В., Тесля Д. В. Запасы углерода в фитомассе древесных растений и крупных древесных остатках в старовозрастных сосняках черничных заповедника «Кивач» // Вопросы лесной науки. 2024. Т. 7. № 4. Статья 156. DOI: 10.31509/2658-607x-202474-156
Приказ Минприроды России № 371 от 27.05.2022 «Об утверждении методик количественного определения объемов выбросов парниковых газов и поглощений парниковых газов» URL: https://clc.li/uOxvY (дата обращения 03.02.2026).
Трофимова И. Л., Кощеева У. П., Нагимов З. Я. Надземная фитомасса сосновых насаждений в различных типах леса в условиях Среднего Урала // АВУ. 2012. № 8 (100). С. 55–58.
Усольцев В. А. Фитомасса модельных деревьев лесообразующих пород Евразии: база данных, климатически обусловленная география, таксационные нормативы. Екатеринбург: изд-во Уральского государственного лесотехнического университета, 2016. 336 с.
Цветков В. Ф., Чертовский В. Г. Классификационные типологические схемы лесов и лесорастительное районирование Мурманской области. Архангельск: Изд-во АИЛиЛХ, 1979. 36 с.
Чепурко Н. Л. Биологическая продуктивность и круговорот химических элементов в лесных и тундровых сообществах Хибинских гор // Биологическая продуктивность и круговорот химических элементов в растительных сообществах. Л.: Наука, 1971. С. 213–219.
Чепурко Н. Л. Структура и годовой баланс биомассы в лесах Хибинских гор / Почвы и продуктивность растительных сообществ. Вып. 1. М.: МГУ, 1972. С. 94–116.
Шешницан С. С., Карташова Н. П., Штепа Е. Н., Царегородцев А. В., Сафонова А. А. Запасы углерода в фитомассе и биологическая продуктивность спелых и перестойных древостоев пригородного лесничества Воронежской области // Лесотехнический журнал. 2024. Т. 14. № 4(56). С. 97–110. DOI: 10.34220/issn.2222-7962/2024.4/7
Экологический атлас Мурманской области / гл. ред. Г. В. Калабин. М.; Апатиты. ИППЭС КНЦ РАН; Геогр. ф-т МГУ; Гос. ком. по охране окружающей среды Мурманской области, 1999. 48 с.
Beck H. E., McVicar T. R., Vergopolan N., Berg A., Lutsko N. J., … & Miralles D. G. High-resolution (1 km) Köppen-Geiger maps for 1901–2099 based on constrained CMIP6 projections // Scientific Data. 2023. Vol. 10. P. 724. DOI: 10.1038/s41597-023-02549-6
Bonan G. B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests // Science. 2008. Vol. 320. P. 1444–1449. DOI: 10.1126/science.1155121
Davidson S. J., Davies M. A., Wegener E., Claussen S., Schmidt M., Peacock M., Strack M. Carbon stocks and fluxes from a boreal conifer swamp: Filling a knowledge gap for understanding the boreal C cycle // Journal of Geophysical Research: Biogeosciences. 2024. Vol. 129. DOI: 10.1029/2024JG008005
Framstad E., de Wit H., Mäkipää R., Larjavaara M., Vesterdal L., Karltun E. Biodiversity, carbon storage and dynamics of old northern forests. Copenhagen: Nordic Council of Ministers, 2013. 130 p. DOI: 10.6027/TN2013-507
Friedlingstein P., O’Sullivan M., Jones M., Andrew R., Hauck J., Landschützer P. … & Zeng J. Global Carbon Budget 2024 // Earth System Science Data. 2025. Vol. 17. P. 965–1039. DOI: 10.5194/essd-17-965-2025
Ķeniņa L., Elferts D., Baders E., Jansons A. Carbon Pools in a Hemiboreal Over-Mature Norway Spruce Stands // Forests. 2018. Vol. 9. No. 7. P. 435. DOI: 10.3390/f9070435
Luyssaert S., Schulze E. D., Börner A., Knohl A., Hessenmöller D., Law B. E., Ciais P., Grace J. Old-growth forests as global carbon sinks // Nature. 2008. Vol. 455. P. 213–215. DOI: 10.1038/nature07276
Merilä P., Lindroos A. J., Helmisaari H S. Hilli S., Nieminen T. M., Nöjd P., Rautio P., Salemaa M., Ťupek B., Ukonmaanaho L. Carbon Stocks and Transfers in Coniferous Boreal Forests Along a Latitudinal Gradient // Ecosystems. 2024. Vol. 27. P. 151–167. DOI: 10.1007/s10021-023-00879-5
Mo L., Zohner C. M., Reich P. B., Liang J., de Miguel S., … & Ortiz-Malavasi E. Integrated global assessment of the natural forest carbon potential // Nature. 2023. Vol. 624. № 7990. P. 92–101. DOI: 10.1038/s41586-023-06723-z
Nunes L. J. R., Meireles C. I. R., Pinto Gomes C. J., Almeida Ribeiro N. M. C. Forest contribution to climate change mitigation: management oriented to carbon capture and storage // Climate. 2020. Vol. 8. No 2. P. 21. DOI: 10.3390/cli8020021
Pan Y., Birdsey R. A., Phillips O. L., Houghton R., Fang J., … & Murdiyarso D. The enduring world forest carbon sink // Nature. 2024. Vol. 631. P. 536–569. DOI: 10.1038/s41586-024-07602-x
Pappas C., Maillet J., Rakowski S., Baltzer J. L., Barr A. G., … & Zha T. Aboveground tree growth is a minor and decoupled fraction of boreal forest carbon input // Agricultural and Forest Meteorology. 2020. Vol. 290. P. 108030. DOI: 10.1016/j.agrformet.2020.108030
Repola J., Ojansuu R., Kukkola M. Biomass Functions for Scots Pine, Norway Spruce and Birch in Finland. Helsinki: Finnish Forest Research Institute, 2007. 28 p.
Schepaschenko D., Moltchanova E., Fedorov S., Karminov V., Ontikov P., … & Kraxner F. Russian forest sequesters substantially more carbon than previously reported // Scientific Reports. 2021. Vol. 11. P. 12825. DOI: 10.1038/s41598-021-92152-9
Strîmbu V. F., Næsset E., Ørka H. O., Liski J., Petersson H., Gobakken T. Estimating biomass and soil carbon change at the level of forest stands using repeated forest surveys assisted by airborne laser scanner data // Carbon Balance Manage. 2023. Vol. 18. No. 10. DOI: 10.1186/s13021-023-00222-4
Thurner M., Beer C., Santoro M., Carvalhais N., Wutzler T., … & Schmullius C. Carbon stock and density of boreal and temperate forests // Global Ecology and Biogeography. 2014. Vol. 23 P. 297–310. DOI: 10.1111/geb.12125
Wirth C., Schumacher J., Schulze E. D. Generic biomass functions for Norway spruce in Central Europe – a meta-analysis approach toward prediction and uncertainty estimation // Tree Physiology. 2004. Vol. 24. No. 2. P. 121–139. DOI: 10.1093/treephys/24.2.121
Zianis D., Muukkonen P., Mäkipää R., Mencuccini M. Biomass and stem volume equations of tree species in Europe // Silva Fennica. 2005. No. 4. DOI: 10.14214/sf.sfm4
Рецензент: к. с.-х. н., в. н. с. Мошников С. А.





