- DOI 10.31509/2658-607x-2020-3-2-1-24
- УДК: 574.3+595.14
Сообщества дождевых червей (Oligochaeta, Lumbricidae) хвойных и мелколиственных лесов лесостепного Приобья
![]() © 2020 г.
© 2020 г.
С.А. Ермолов
Новосибирский национальный исследовательский государственный университет, Россия, 630090 Новосибирск, ул. Пирогова, 1
E-mail: ermserg96@gmail.com
Поступила в редакцию 06.04.2020 г.
Принята к печати 02.06.2020 г.
Дождевые черви – значимая группа почвенной макрофауны, обеспечивающая ряд экосистемных функций в лесах. Исследования видового состава и плотности населения дождевых червей проводились во многих регионах России, тем не менее, люмбрикофауна Новосибирской области долгое время оставалась неизученной. Цель данной работы – сравнительный анализ населения дождевых червей хвойных и мелколиственных лесов лесостепного Приобья Новосибирской области и выявление взаимосвязей его состава с основными физико-химическими свойствами почвы. Исследование проведено в сосновых борах и березово-осиновых лесах. В качестве основного метода учета использовалась послойная раскопка с ручным разбором почвенных проб и разбор лесного валежника. Также измерены некоторые почвенные характеристики. Приведены данные по видовому составу и плотности населения дождевых червей для каждого местообитания. Составлена классификация исследованных местообитаний в зависимости от соотношения жизненных форм дождевых червей. Установлено, что влажность почвы наиболее значимый фактор для группы поверхностнообитающих видов. Разнообразие поверхностнообитающих видов дождевых червей в лесах во многом поддерживается за счет валежника. Для азиатского подвида Eisenia nordenskioldi nordenskioldi проведен морфометрический анализ, подтверждающий характерный для него полиморфизм.
Ключевые слова: дождевые черви, жизненные формы, Eisenia nordenskioldi nordenskioldi, Новосибирская область, сосновые боры, мелколиственные леса, валежник, свойства почв
В функционировании лесных экосистем дождевые черви выступают одной из важных групп организмов-почвообразователей. Будучи сапрофагами, дождевые черви разных видов и жизненных форм в разной степени обеспечивают трансформацию органического вещества: подстилочные и почвенно-подстилочные черви разлагают растительные останки, тем самым участвуя в формировании гумусового горизонта; норные черви обеспечивают проникновение органики в глубокие слои почвы; собственно-почвенные черви, обитающие на разной глубине, потребляют гумус, осуществляя процессы минерализации органики и перемещения соединений C и N в почве (Перель, 1975; 1979; Holdsworth et al., 2008). Присутствие тех или иных видов и жизненных форм дождевых червей, а также оценка их плотности населения служат индикаторами состояния лесных почв (Чекановская, 1960; Аккумуляция…, 2018).
Почва является далеко не единственным местообитанием дождевых червей. Они также населяют лесную подстилку, живут в скоплениях экскрементов животных, обильно встречаются в валежнике, участвуя в его деструкции. Последнее наиболее важно для лесных экосистем, где не только видовое, но и функциональное разнообразие дождевых червей может поддерживаться за счет валежника (Гераськина, 2016б, 2016в; Ashwood et al., 2019).
Исследования дождевых червей в лесных экосистемах проводились во многих регионах России. Более подробно были изучены территория Русской равнины, Урал, Северо-Западный и Центральный Кавказ, Дальний Восток (Перель, 1958; Пенев и др., 1994; Шашков, 2003; Рапопорт, 2010; Ганин, 2013). В Западной Сибири исследовались леса Алтая и Горной Шории (Перель, 1994; Мусиенко, 2019), а на территории Новосибирской области, в частности в районе лесостепного Приобья, для которого характерны разнообразные лесные ландшафты (Мугако, 2008; Атлас…, 2002), подобные исследования практически не проводились. Поскольку в этом регионе с недавнего времени начались работы по изучению биотопического распределения и молекулярно-генетического разнообразия дождевых червей (Шеховцов и др., 2016; Ким-Кашменская, 2016; Ермолов, 2018а, 2018б, 2019; Шеховцов, Ермолов и др., 2020), нами было решено продолжить исследования, изучив лесные экосистемы лесостепного Приобья Новосибирской области..
Цель данной работы – провести сравнительный анализ населения дождевых червей хвойных и мелколиственных лесов лесостепного Приобья Новосибирской области и выявить взаимосвязь его состава с основными факторами среды обитания.
Задачи: 1. Сравнить видовой состав и комплексы жизненных форм дождевых червей (Lumbricidae) хвойных (сосновые боры) и мелколиственных (березово-осиновые) лесов.
- Изучить население дождевых червей лесного валежника, как специфического местообитания почвенных сапрофагов.
- Исследовать взаимосвязи видового состава дождевых червей с основными физико-химическими свойствами лесных почв.
- Обосновать выделение крупной и мелкой размерных форм подвида Eisenia nordenskioldi nordenskioldi по внешним морфологическим признакам.
МАТЕРИАЛ И МЕТОДЫ
Регион исследований. Новосибирская область традиционно подразделяется на пять природно-географических регионов: Васюганье, Бараба, Кулунда, Приобье и Салаир (Мугако, 2008; Атлас…, 2002). Территория лесостепного Приобья включает в себя как типичные лугово-степные ландшафты, так и разнотравные боры и мелколиственные (березово-осиновые) травяные леса на дерново-подзолистых и серых лесных почвах (Атлас…, 2002), в которых был собран материал для исследования. Учеты проведены в высокотравно-папоротниковых борах (Заельцовский, Кудря-шовский, Чемской, боры в окрестностях села Барышево и поселка Сосновка); в березово-осиновых лесах: ввысокотравно-папоротниковые березово-осиновые леса (окрестности села Быково, поселков Морозово и Кольцово), высокотравно-снытиевый березовый лес (окрестности станции Шелковичиха), низкотравный березовый лес (окрестности села Верх-Тула). На данных территориях отмечены следующие типы почв: дерново-слабоподзолистые песчаные (боровые пески), дерново-подзолистые и серые лесные (в основном, темные) (рис. 1) (Почвенная карта…, 2007).
Сбор дождевых червей. Учет дождевых червей в почве проводился с использованием послойной раскопки и ручного разбора проб (Методы…, 1975). На выбранном участке лесной почвы размечался квадрат 50 см. Вначале перебиралась подстилка, затем выкапывались слои почвы толщиной 0-2 см, 2-5 см, 5-10 см и более 10 см (~до 40 см или глубже) с последующим ручным разбором. В каждом исследованном лесу взято по 8 почвенных проб.
Учёт дождевых червей в валежнике: у обнаруженных упавших стволов 2-3-й стадий разложения или их фрагментов измерялись длина и диаметр в нескольких местах, а затем проводился ручной разбор коры, моховых наростов и гниющей древесины. По возможности, валежник разбирался до почвы (Гераськина, 2016б, 2016в).
Собранные черви умерщвлены в 2% растворе формальдегида и зафиксированы в 4% растворе формальдегида с глице-рином (Чекановская, 1960). Видовой состав дождевых червей был установлен по определителю Т.С. Всеволодовой-Перель (Всеволодова-Перель, 1997).
Для особей подвида Eisenia nordenskioldi nordenskioldi, с характерным ярко выраженным полиморфизмом (Перель, 1994, 1997), был проведен морфометрический анализ с измерением следующих признаков (Шеховцов, Ермолов и др., 2020): количество сегментов (КС), длина тела (Д, мм), ширина тела в самом широком месте (Ш, мм), длина пояска (ДП, мм), ширина пояска (ШП, мм), вес зафиксированного в растворе формальдегида червя (В, г). Поскольку, черви крупной размерной формы, пригодные для морфометрии (то есть взрослые черви с отчётливо выраженным пояском) встречались редко, то из всех найденных экземпляров в борах и мелколиственных лесах были составлены две общие выборки. Выборки червей мелкой размерной формы составлены для конкретных местообитаний: две для валежников (Кольцово, Быково) и одна для почвы (Верх-Тула), где численность подвида была наиболее высока.
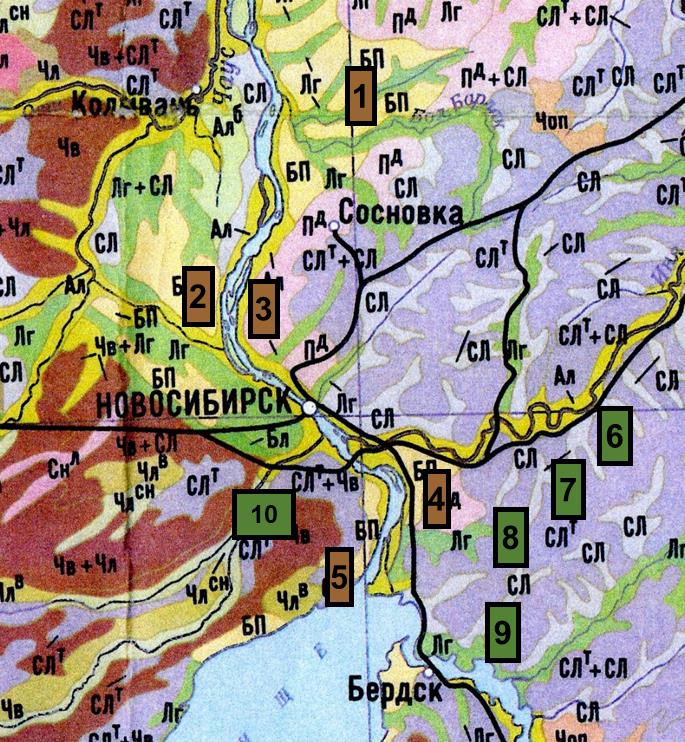
Рисунок 1. Фрагмент почвенной карты Новосибирской области с исследованными борами и мелколиственными лесами
Условные обозначения: Сосновые боры: 1 – окрестности п. Сосновка; 2 – Кудряшовский бор; 3 – Заельцовский бор; 4 – окрестности с. Барышево; 5 – Чемской бор; Мелколиственные леса в окрестностях населенных пунктов: 6 – ст. Шелковичиха; 7 – с. Быково; 8 – п. Кольцово; 9 – п. Морозово; 10 – с. Верх-Тула
Учеты проведены летом 2019 года. Всего за сезон было взято 80 почвенных проб, разобрано 5.13 м3 валежника и собрано 3394 особей дождевых червей разных видов и жизненных форм.
Анализ почвенных проб. Во время сбора дождевых червей во всех исследованных местообитаниях для каждой почвенной пробы проводились измерения мощности подстилки и гумусового горизонта (с точностью до 1 мм); из каждого слоя пробы (0-2, 2-5, 5-10, 10-30 см) взяты образцы почвы для дальнейшего лабораторного анализа.
В почвенных образцах определены влажность (гигроскопическая влажность) и кислотность почвы (актуальная кислотность, рН водн.). Для расчёта содержания влаги в почве свежие влажные пробы были расфасованы в алюминиевые стаканчики и взвешены, затем доведены до воздушно-сухого состояния и повторно взвешены. Доля влаги (W) в процентах рассчитывалась по формуле:
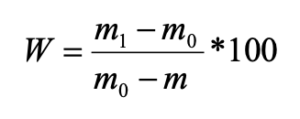
Где m1 – масса важной почвы со стаканчиком и крышкой, г; m0 – масса высушенной почвы со стаканчиком и крышкой, г; m – масса пустого стаканчика с крышкой, г (ГОСТ 28268-89).
Кислотность измеряли потенциомет-рическим методом: из 20 г. навески сухой почвы, просеянной на мелком сите, готовили суспензию в дистиллированной воде, а затем помещали в неё электрод pH-метра МИ-150 (Воробьева и др., 2012).
Обработка данных. Полученные данные о плотности населения дождевых червей были рассчитаны на единицу площади для почвенных проб (особей/м2), и на единицу объёма (особей/м3) для валежника; объём валежника вычислялся по формуле: V=π*r²*h, где r – средний радиус ствола; h – высота ствола (Гераськина, 2016б, 2016в).
Для расчета корреляций между плотностью населения дождевых червей и физико-химическими свойствами почвы был использован коэффициент корреляции Спирмена.
Для каждого местообитания были рассчитаны средние показатели данных по влажности и кислотности почвы. Средние значения также рассчитаны для мощности подстилки и гумусового горизонта. Далее было проведено сопоставление полученных данных с плотностью населения дождевых червей в каждой группе местообитаний. Отдельно рассматривались черви, относящиеся к основным морфо-экологическим типам (Перель, 1975, 1979): черви, питающиеся на поверхности почвы (подстилочные и почвенно-подстилочные) и черви, питающиеся почвенным гумусом (собственно-почвенные верхне- и среднеярусные). В корреляционном анализе не учитывались данные по плотности собственно-почвенных червей в бору в окрестностях села Барышево – в связи с низкой плотностью червей этой группы (10 особей/м2).
Выборки с морфометрическими данными дождевых червей E. nordenskioldi nordenskioldi сравнивались с помощью критерия Уитни-Манна.
Выполнена кластеризация местообитаний по составу жизненных форм в программе Past. Исходные данные нормированы по показателю доминации: во всех местообитаниях плотность населения дождевых червей каждой жизненной формы рассчитывалась как доля от общего населения. Это позволило получить однородные единицы измерения. Для измерения расстояния между объектами было использовано Евклидово расстояние, а для построения дендрограмм использовался метод Варда (Уорда) (Вэн Райзин, 1980).
РЕЗУЛЬТАТЫ
Население дождевых червей в сосновых борах. В исследованных сосновых борах отмечены как космополитные виды дождевых червей (Dendrobaena octaedra (Savigny, 1826), Lumbricus rubellus Hoffmeister, 1843, Aporrectodea caliginosa caliginosa (Savigny, 1826), Dendrodrilus rubidus subrubicundus (Eisen, 1874), так и черви, населяющие в основном азиатскую часть РФ и сопредельные территории (Eisenia nordenskioldi nordenskioldi (Eisen, 1879), Eisenia nordenskioldi pallida Malevič, 1956).
Наибольшая плотность дождевых червей отмечена в Заельцовском бору (174 особи/м2), а наименьшая – в бору в окрестностях села Барышево (30 особей/м2), где черви были обнаружены не во всех пробах, а некоторые виды встречались единично (табл. 1). Наиболее разнообразный видовой состав обнаружен в Кудряшовском бору (4 подвида, 2 вида; всего 172 особи/м2, некоторые виды единичны), а наименее разнообразный – в бору в окрестностях поселка Сосновка (2 подвида, 1 вид; всего 124 особи/м2), где, за исключением D. octaedra, отсутствовали космополитные виды. Чемской бор схож по видовому составу с Кудряшовским, а по значению плотности населения червей с бором в поселке Сосновка (3 подвида, 2 вида; всего 124 особи/м2). Во всех исследованных борах подвид E. n. nordenskioldi представлен как мелкой, так и крупной размерными формами.
Таблица 1. Население дождевых червей (Lumbricidae) в борах (X ± SE)
| Тип биотопа | Место сбора | Жизненные формы | Вид | Плотность населения вида, особи/м2 | Доля жизненной формы, % |
| Высокотравно-папоротниковые боры | Чемской бор | Подстилочные | D. octaedra | 4±2 | 3 |
| Почвенно-подстилочные | L. rubellus | 2±1 | 11 | ||
| E. n. nordenskioldi (мелкая форма) | 6±2 | ||||
| E. n. nordenskioldi (крупная форма) | 6±2 | ||||
| Собственно-почвенные (среднеярусные) | A. c. caliginosa | 84±23 | 86 | ||
| E. n. pallida | 22±8 | ||||
| Кудряшовский бор | Подстилочные | D. octaedra | 16±3 | 11 | |
| D. r. subrubicundus | 3±1 | ||||
| Почвенно-подстилочные | Lumbricus rubellus | 1±1 | 10 | ||
| E. n. nordenskioldi (мелкая форма) | 11±3 | ||||
| E. n. nordenskioldi (крупная форма) | 5±2 | ||||
| Собственно-почвенные (среднеярусные) | A. c. caliginosa | 80±33 | 79 | ||
| Eisenia nordenskioldi pallida | 57±20 | ||||
| Заельцовский бор | Подстилочные | Dendrobaena octaedra | 14±8 | 8 | |
| Почвенно-подстилочные | Lumbricus rubellus | 42±16 | 32 | ||
| E. n. nordenskioldi (мелкая форма) | 10±4 | ||||
| E. n. nordenskioldi (крупная форма) | 4±1 | ||||
| Собственно-почвенные (среднеярусные) | A. c. caliginosa | 42±26 | 60 | ||
| E. n. pallida | 62±20 | ||||
| Бор в окрестностях поселка Сосновка | Подстилочные | D. octaedra | 16±7 | 13 | |
| Почвенно-подстилочные | E. n. nordenskioldi (мелкая форма) | 32±17 | 35 | ||
| E. n. nordenskioldi (крупная форма) | 12±8 | ||||
| Собственно-почвенные (среднеярусные) | E. n. pallida | 64±43 | 52 | ||
| Бор в окрестностях села Барышево | Подстилочные | D. octaedra | 9±3 | 34 | |
| D. r. subrubicundus | 1±1 | ||||
| Почвенно-подстилочные | E. n. nordenskioldi (мелкая форма) | 9±3 | 33 | ||
| E. n. nordenskioldi (крупная форма) | 1±1 | ||||
| Собственно-почвенные (верхнеярусные) | O. lacteum (мелкая форма) | 4±4 | 13 | ||
| Собственно-почвенные (среднеярусные) | E. n. pallida | 6±3 | 20 |
По соотношению жизненных форм все исследованные боры разделились на три группы:
- боры с преобладанием (до 86%), собственно-почвенных, среднеярусных форм (Чемской и Кудряшовский боры);
- боры, где около 50% населения составляют поверхностнообитающие черви
(Заельцовский бор и окрестности поселка Сосновка);
- бор, в котором соотношение всех жизненных форм примерно одинаково (окрестности села Барышево) (рис. 2; табл. 1).
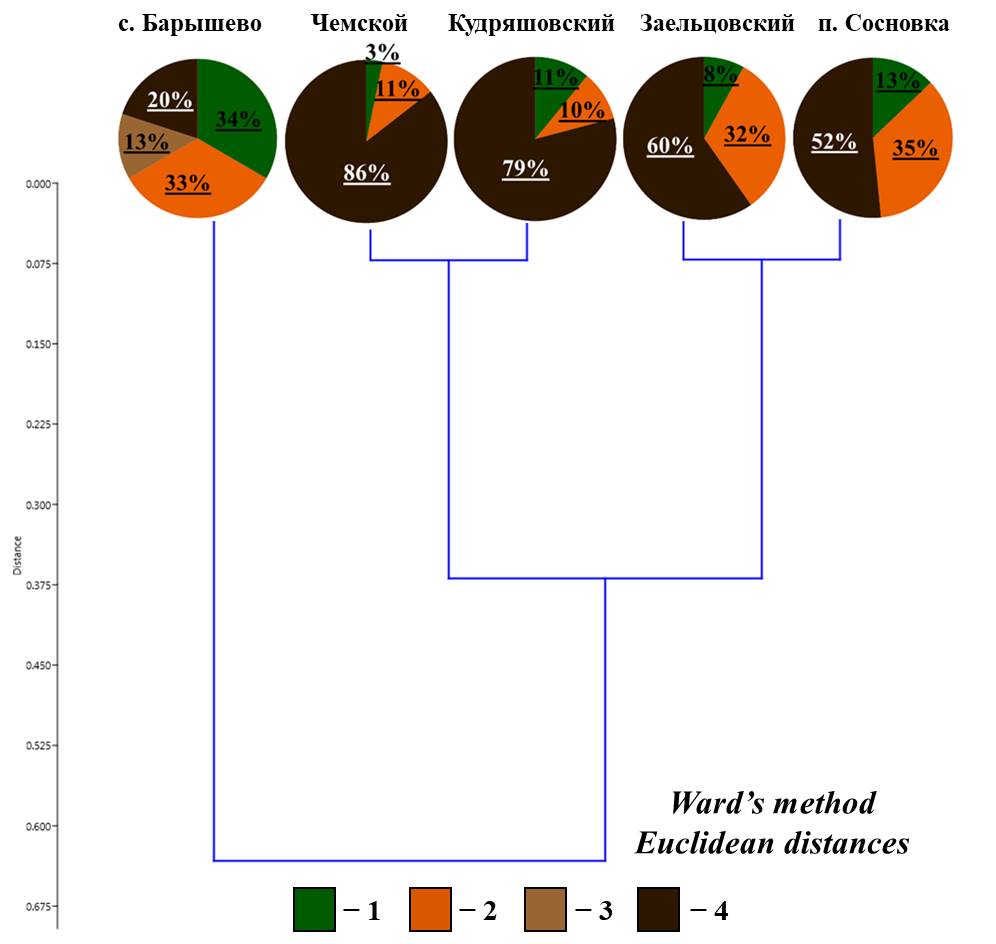
Рисунок 2. Соотношение жизненных форм дождевых червей в сосновых борах
Условные обозначения: 1 – подстилочные; 2 – почвенно-подстилочные; 3 – собственно-почвенные верхнеярусные; 4 – собственно-почвенные среднеярусные
При этом в Чемском и Кудряшовском борах видовой состав, формирующий комплексы жизненных форм практически одинаков, а в Заельцовском бору и окрестностях п. Сосновка выявлены значительные различия в видовом составе. Так, почвенно-подстилочные формы в Заельцовском бору представлены преимущественно видом L. rubellus, а в Сосновке из этой группы обитает E. n. nordenskioldi. В Заельцовском бору, собственно-почвенные, среднеярусные формы представлены подвидами A. c. caliginosa и E. n. pallida (при этом, плотность населения A. c. caliginosa заметно ниже, чем в Чемском и Кудряшовском борах), а в Сосновке данную форму представляет только E. n. pallida. В бору в окрестностях села Барышево единично отмечен собственно-почвенный верхнеярусный вид Octolasion lacteum (Örley, 1885), а другие виды и жизненных формы обнаружены примерно в равном соотношении.
Население дождевых червей в мелколиственных лесах. В исследованных мелколиственных лесах видовой состав дождевых червей в целом был схож с таковым в борах (табл. 2). Более высокие показатели плотности населения дождевых червей отмечены в двух высокотравно-папоротниковых березово-осиновых лесах (окрестности села Быково — 216 особей/м2 и поселка Морозово — 263 особей/м2) и высоктравно-снытиевом березовом лесу (окрестности станции Шелковичиха — 267 особей/м2). Также в этих лесах обнаружен вид O. lacteum (мелкая форма), плотность населения которого была наибольшей в лесу в окрестностях поселка Морозово (202 особей/м2).
В высокотравно-папоротниковом березово-осиновом лесу в окрестностях поселка Кольцово и в низкотравном березовом лесу в окрестностях села Верх-Тула плотность населения червей была значительно ниже (Кольцово — 55 особей/м2, Верх-Тула — 72 особей/м2). Примечательно и небольшое видовое разнообразие – только три вида: E. n. nordenskioldi, E. n. pallida и единично D. octaedra. Также низкотравный березовый лес (Верх-Тула) единственный из рассмотренных биотопов, где не обнаружена крупная форма E. n. nordenskioldi, но при этом плотность мелкой формы сравнительно высока (табл. 2).
На полученной дендрограмме соотношения жизненных форм (рис. 3) все мелколиственные леса разделяются на две большие группы:
- в лесу в окрестностях поселка Морозово преобладающей жизненной формой (88% численности) являются, собственно-почвенные, верхнеярусные черви (O. lacteum);
- во вторую группу вошли остальные леса, где доля собственно-почвенных среднеярусных червей составляет около половины от общего населения. Внутри этой группы есть различия: в трех лесах значительно представлены поверхностно-обитающие черви (окрестности поселка Кольцова и села Верх-Тула – почвенно-подстилочные, станции Шелковичихи – подстилочные), а в лесу в окрестностях с. Быково 33% населения составляют собственно-почвенные верхнеярусные черви. Также, следует отметить, что почвенно-подстилочные черви в лесах окрестностей Кольцово и Верх-Тулы представлены только n. nordenskioldi, а собственно-почвенные, среднеярусные E. n. pallida. В лесах окрестностей села Быково и станции Шелковичиха, оба подвида также встречаются, однако основную долю двух жизненных форм составляют L. rubellus и A. c. caliginosa соответственно (табл. 2).
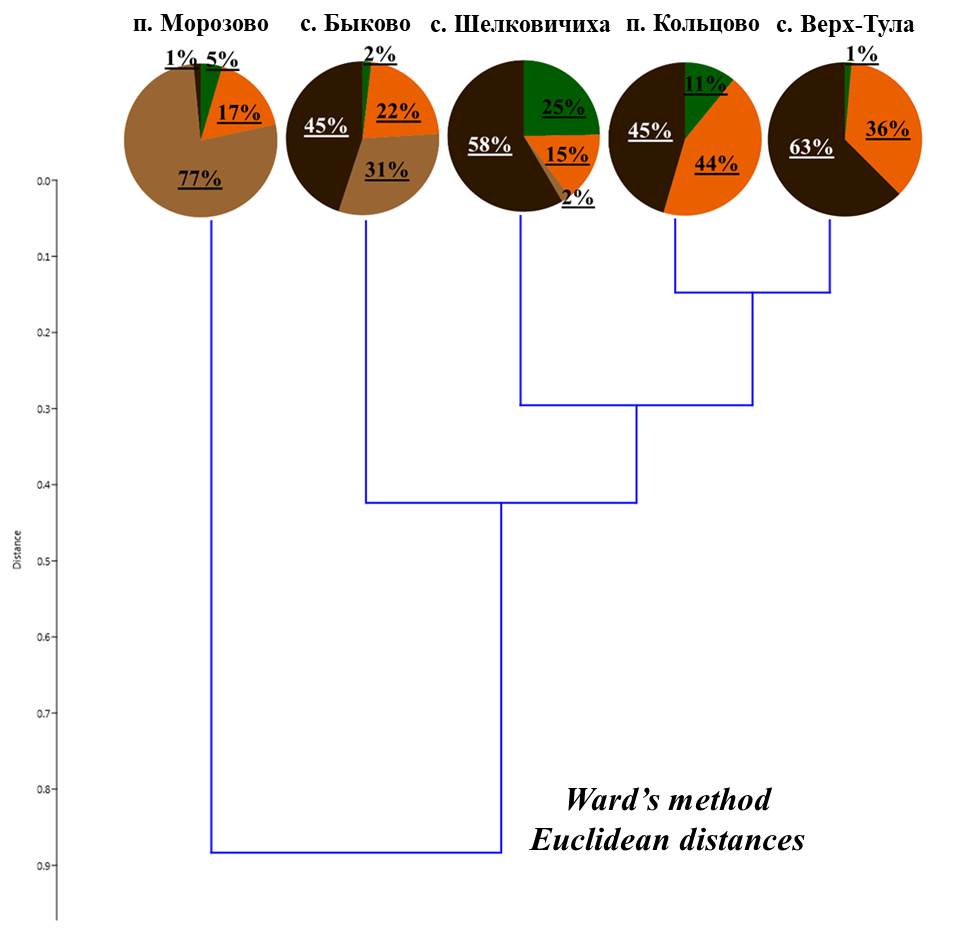
Рисунок 3. Соотношение жизненных форм дождевых червей в мелколиственных лесах
Примечание: здесь и далее условные обозначения как для рис. 2
Таблица 2. Население дождевых червей (Lumbricidae) в мелколиственных лесах (X ± SE)
| Тип биотопа | Место сбора | Жизненные формы | Вид | Плотность населения, особи/м2 | Доля жизненной формы, % |
| Высокотравно-папоротниковые березово-осиновые леса | Окрестности с. Быково | Подстилочные | D. octaedra | 5±2 | 2 |
| Почвенно-подстилочные | L. rubellus | 39±14 | 22 | ||
| E. n. nordenskioldi
(мелкая форма) |
8±3 | ||||
| E. n. nordenskioldi
(крупная форма) |
1±1 | ||||
| Собственно-почвенные (верхнеярусные) | O. lacteum
(мелкая форма) |
67±21 | 31 | ||
| Собственно-почвенные (среднеярусные) | A. c. caliginosa | 95±34 | 45 | ||
| E. n. pallida | 2±2 | ||||
| Окрестности п. Морозово | Подстилочные | D. octaedra | 12±3 | 5 | |
| Почвенно-подстилочные | E. n. nordenskioldi
(мелкая форма) |
39±8 | 17 | ||
| E. n. nordenskioldi
(крупная форма) |
6±2 | ||||
| Собственно-почвенные (верхнеярусные) | O. lacteum
(мелкая форма) |
202±29 | 77 | ||
| Собственно-почвенные (среднеярусные) | A. c. caliginosa | 4±2 | 1 | ||
| Окрестности п. Кольцово | Подстилочные | D. octaedra | 6±3 | 11 | |
| Почвенно-подстилочные | E. n. nordenskioldi
(мелкая форма) |
23±6 | 44 | ||
| E. n. nordenskioldi
(крупная форма) |
1±1 | ||||
| Собственно-почвенные (среднеярусные) | E. n. pallida | 25±6 | 45 | ||
| Высокотравно-снытиевый березовый лес | Окрест. ст. Шелковичиха | Подстилочные | D. octaedra | 64±16 | 25 |
| D. r. tenuis | 3±2 | ||||
| Почвенно-подстилочные | L. rubellus | 27±12 | 15 | ||
| E. n. nordenskioldi
(мелкая форма) |
8±4 | ||||
| E. n. nordenskioldi
(крупная форма) |
5±2 | ||||
| Собственно-почвенные (верхнеярусные) | O. lacteum
(мелкая форма) |
5±4 | 2 | ||
| Собственно-почвенные (среднеярусные) | A. c. caliginosa | 146±48 | 58 | ||
| E. n. pallida | 10±8 | ||||
| Низкотравный березовый лес | Окрестности с. Верх-Тула | Подстилочные | D. octaedra | 1±1 | 1 |
| Почвенно-подстилочные | E. n. nordenskioldi
(мелкая форма) |
26±5 | 36 | ||
| Собственно-почвенные (среднеярусные) | E. n. pallida | 45±13 | 63 |
Население дождевых червей в лесном валежнике. Валежник, подходящий для учета дождевых червей (Гераськина, 2016б, 2016в) был обнаружен только в пяти местообитаниях: в трех борах (Заельцовский, Чемской, окрестности села Бырашево) и в двух мелколиственных лесах (высокотравно-папоротниковые березово-осиновые леса в с Быково и поселка Кольцово). Исследованные валежники представляли собой одиночные упавшие стволы Pinus sylvestris и Betula pendula разного размера 2-3-й стадий разложения. Практически все население валежников составляли подстилочные и почвенно-подстилочные черви, лишь изредка в стволах 3-й стадии разложения встречались собственно-почвенные черви (табл. 3, табл. 4). Примечательно, что в бору в окрестностях села Барышево и в березово-осиновом лесу в окрестностях поселка Кольцово, где плотность населения дождевых червей в почве была низкой, валежники отличались высокой плотностью населения и разнообразием видового состава.
Таблица 3. Население дождевых червей (Lumbricidae) в валежниках боров
| Место сбора | Стадия разложения, порода дерева | Жизненные формы | Виды | Плотность населения, особи/м3 | Доля жизненной формы, % | |
| Высокотравно-папоротниковые боры | ||||||
| Заельцовский бор | 2, сосна | Подстилочные | D. octaedra | 101 | 88 | |
| D. r. tenuis | 8 | |||||
| Почвенно-подстилочные | L. rubellus | 14 | 12 | |||
| E. n. nordenskioldi (мелкая форма) | 1 | |||||
| Чемской бор | 2, береза | Подстилочные | D. octaedra | 64 | 54 | |
| D. r. tenuis | 48 | |||||
| Почвенно-подстилочные | E. n. nordenskioldi (мелкая форма) | 84 | 40 | |||
| Собственно-почвенные (среднеярусные) | E. n. pallida | 12 | 6 | |||
| Бор в окрестностях села Барышево | 2, сосна | Подстилочные | D. octaedra | 192 | 99 | |
| D. r. tenuis | 169 | |||||
| Почвенно-подстилочные | E. n. nordenskioldi (мелкая форма) | 4 | 1 | |||
| 3, сосна | Подстилочные | D. octaedra | 12 | 44 | ||
| D. r. tenuis | 13 | |||||
| Почвенно-подстилочные | E. n. nordenskioldi (мелкая форма) | 14 | 25 | |||
| Собственно-почвенные (верхнеярусные) | O. lacteum (мелкая форма) | 17 | 29 | |||
| Собственно-почвенные (среднеярусные) | E. n. pallida | 1 | 2 | |||
| 3, береза | Подстилочные | D. octaedra | 164 | 88 | ||
| D. r. tenuis | 120 | |||||
| Почвенно-подстилочные | E. n. nordenskioldi (мелкая форма) | 40 | 12 | |||
Таблица 4. Население дождевых червей (Lumbricidae) в валежниках мелколиственных лесов
| Место сбора | Стадия разложения, порода дерева | Жизненные формы | Виды | Плотность населения,
особи/м3 |
Доля жизненной формы, % | |
| Высокотравно-папоротниковые березово-осиновые леса | ||||||
| Окрестности с. Быково | 2, береза №1 | Подстилочные | D. octaedra | 500 | 81 | |
| D. r. tenuis | 600 | |||||
| Почвенно-подстилочные | L. rubellus | 20 | 19 | |||
| E. n. nordenskioldi (мелкая форма) | 240 | |||||
| 2, береза №2 | Подстилочные | D. r. tenuis | 25 | 5 | ||
| Почвенно-подстилочные | L. rubellus | 100 | 95 | |||
| E. n. nordenskioldi (мелкая форма) | 350 | |||||
| 3, береза | Подстилочные | D. r. tenuis | 7 | 22 | ||
| Почвенно-подстилочные | L. rubellus | 6 | 77 | |||
| E. n. nordenskioldi (мелкая форма) | 16 | |||||
| E. fetida | 2 | |||||
| Собственно-почвенные (среднеярусные) | A. c. caliginosa | 1 | 1 | |||
| Окрестности п. Кольцово | 2, береза | Подстилочные | D. octaedra | 4 | 30 | |
| D. r. tenuis | 50 | |||||
| D. r. subrubicundus | 62 | |||||
| Почвенно-подстилочные | E. n. nordenskioldi (мелкая форма) | 273 | 70 | |||
Видовой состав дождевых червей в валежниках Чемского и Заельцовского боров, и березово-осинового леса в окрестностях села Быково сходен с таковым в почве этих местообитаний, за исключением подвида D. r. tenuis (Eisen, 1874), который массово встречается преимущественно в валежнике (Гераськина, 2016г; Ермолов, 2018а). Также валежник в лесу окрестностей села Быково был единственным местообитанием, где обнаружен почвенно-подстилочный вид Eisenia fetida (Savigny, 1896), который в Новосибирской области является синантропным (Ермолов, 2018а, 2018б, 2019).
Для валежников также была проведена классификация по соотношению жизненных форм дождевых червей (рис. 4). Валежники разделились на три группы:
- валежники с преобладанием подстилочных форм;
- валежники с преобладанием почвенно-подстилочных форм;
- валежники с присутствием собственно-почвенных форм червей. Несмотря на сходное соотношение жизненных форм, валежники в пределах группы различаются по преобладанию разных видов дождевых червей, относящихся к одной жизненной форме. Например, в первой группе валежник Заельцовского бора имеет одинаковое соотношение жизненных форм с таковым в окрестностях села Барышево (рис. 4). Но в Заельцовском бору практически все подстилочные черви относятся к виду D. octaedra, почвенно-подстилочные – к виду L. rubellus, а в бору окрестностях Барышево (как, собственно, и в других валежниках этой группы) подстилочные виды представлены D. octaedra и D. r. tenuis, плотности населения которых соотносятся почти как 1:1 (табл. 3, табл. 4), а большую часть почвенно-подстилочных форм составляет подвид n. nordenskioldi (мелкая форма). В группе с доминированием почвенно-подстилочных форм валежники в окрестностях Быково, населяли три разных представителя этой жизненной формы (L. rubellus, E. fetida, E. n. nordenskioldi (мелкая форма), а в окрестностях Кольцово она была представлена только E. n. nordenskioldi (мелкая форма); также только здесь массово встречался подстилочный подвид D. r. subrubicundus. В сильно разложившемся сосновом стволе в бору в окрестностях Барышево обнаружены собственно-почвенные верхнеярусные черви O. lacteum, которые единично встречались в почве; в валежнике Чемского бора отмечена сравнительно высокая доля собственно-почвенных среднеярусных червей E. n. pallida; резкого доминирования среди остальных форм в валежниках этой группы нет.
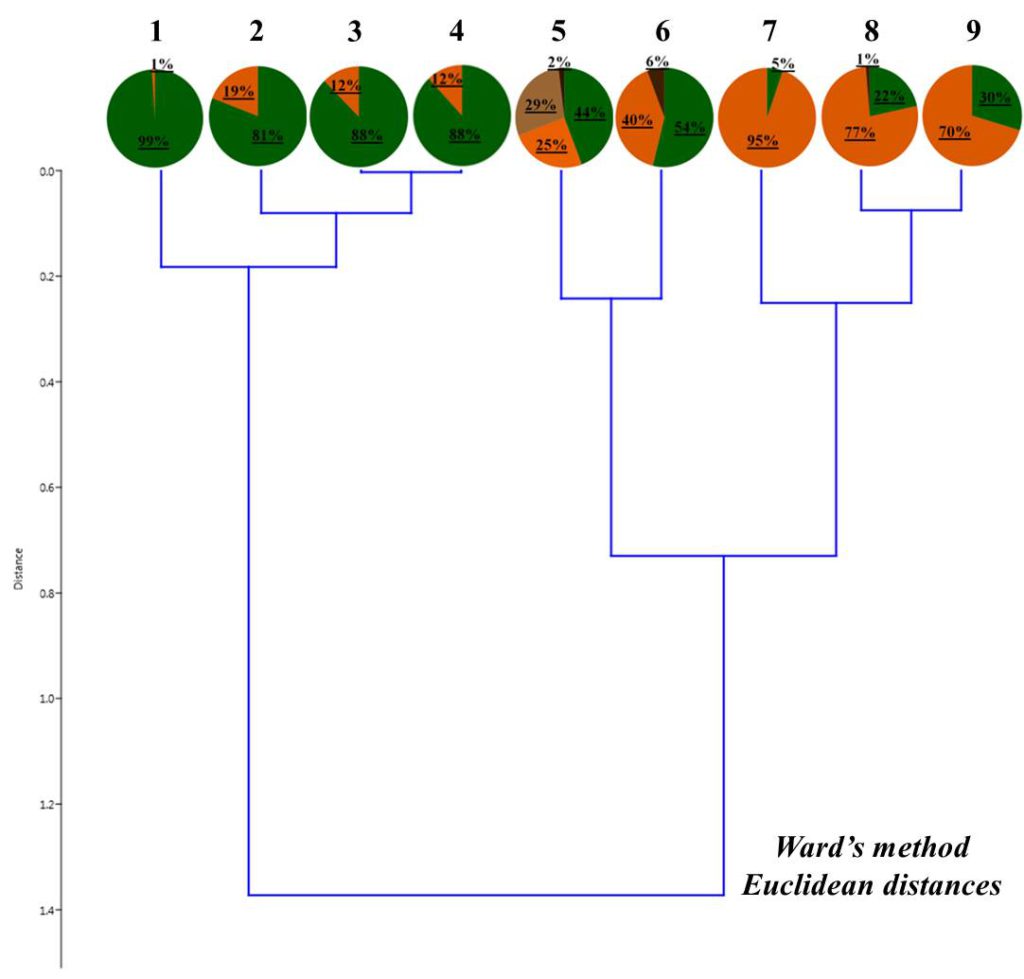
Рисунок 4. Соотношение жизненных форм дождевых червей в валежнике
Условные обозначения: 1– Барышево (сосна ст. 2); 2 – Быково (береза 1 ст. 2); 3 – Заельцовский бор (сосна, ст. 2);
4 – Барышево (береза, ст. 3); 5- Барышево (сосна, ст. 3); 6 – Чемской бор (береза, ст. 2);
7 – Быково (береза 2, ст. 2); 8 – Быково (береза ст. 3); 9 – Кольцово (береза, ст. 2)
Взаимосвязь свойств почвы и видового состава дождевых червей. Измеренные физико-химические характе-ристики почв в исследованных борах и мелколиственных лесах приведены ниже (табл. 5, 6).
Таблица 5. Физико-химические свойства почв сосновых боров (X ± SE)
| Место учета | Мощности подстилки, см | Мощность гумусового горизонта, см | Содержание влаги в почве, % | Кислотность почвы, pH водн. |
| Чемской бор | 3.3±0.1 | 6.8±0.4 | 20.06±1.88 | 5.81±0.08 |
| Заельцовский бор | 2.3±0.2 | 10.4±0.5 | 30.44±3.21 | 5.72±0.06 |
| Кудряшовский бор | 2.4±0.1 | 9.9±0.7 | 25.25±2.22 | 5.38±0.04 |
| Бор в окрестностях поселка Сосновка | 3.5±0.3 | 5±0.4 | 34.81±3.65 | 5.36±0.05 |
| Бор в окрестностях села Барышево | 3.6±0.2 | 10.5±0.7 | 22.5±2.87 | 5.96±0.07 |
Таблица 6. Физико-химические свойства почв мелколиственных лесов (X ± SE)
| Название биотопа | Мощности подстилки, см | Мощность гумусового горизонта, см | Содержание влаги в почве, % | Кислотность почвы, pH водн. |
| Окрестности с. Быково | 1.6±0.2 | 9.9±1.1 | 28±2.24 | 5.68±0.09 |
| Окрестности п. Морозово | 2±0.2 | 19.9±1.1 | 21.5±0.86 | 6.22±0.08 |
| Окрестности п. Кольцово | 2.8±0.2 | 10.3±0.6 | 21.06±0.88 | 6.23±0.06 |
| Окрестности ст. Шелковичиха | 2.2±0.1 | 12.3±0.5 | 39.19±1.63 | 5.42±0.08 |
| Окрестности с. Верх-Тула | 3.4±0.2 | 9.8±0.7 | 17.56±0.75 | 6.35±0.08 |
На диаграмме для боров показано, что наиболее значимым фактором, влияющим на плотность населения поверхностно-обитающих дождевых червей, является влажность почвы (рис. 5). Плотность населения значимо коррелирует с показателями влажности почвы (rs=0.89; p <0.05; n=5). Эти группы обитают у самой поверхности почвы и поэтому более чувствительны к пересыханию. В данных местообитаниях влага задерживается, в основном, за счет подстилки, так как дерново-подзолистые почвы и боровые пески, характерные для боров, слабо удерживают влагу, необходимую дождевым червям для нормальной жизнедеятельности. С другими характеристиками у этой группы взаимосвязи не выявлены.
Для группы собственно-почвенных видов также не установлены взаимосвязи с другими характеристиками почв, кроме влажности. Выявлена положительная связь между плотностью населения E. n. pallida и влажностью почвы (rs=0.90; p <0.05, n=4) и отрицательная между плотностью населения A. c. caliginosa и влажностью почвы (rs=−0.90; p <0.05; n=4) (рис. 6).
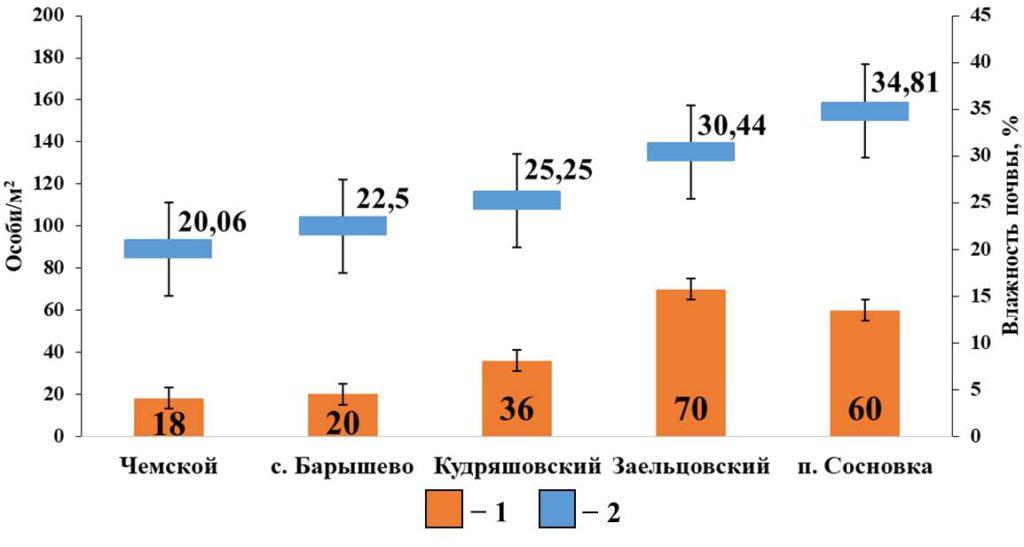
Рисунок 5. Зависимость плотности населения поверхностнообитающих дождевых червей от влажности почвы в сосновых борах
Условные обозначения: 1 — Плотность населения поверхностнообитающих червей; 2 – Влажность почвы
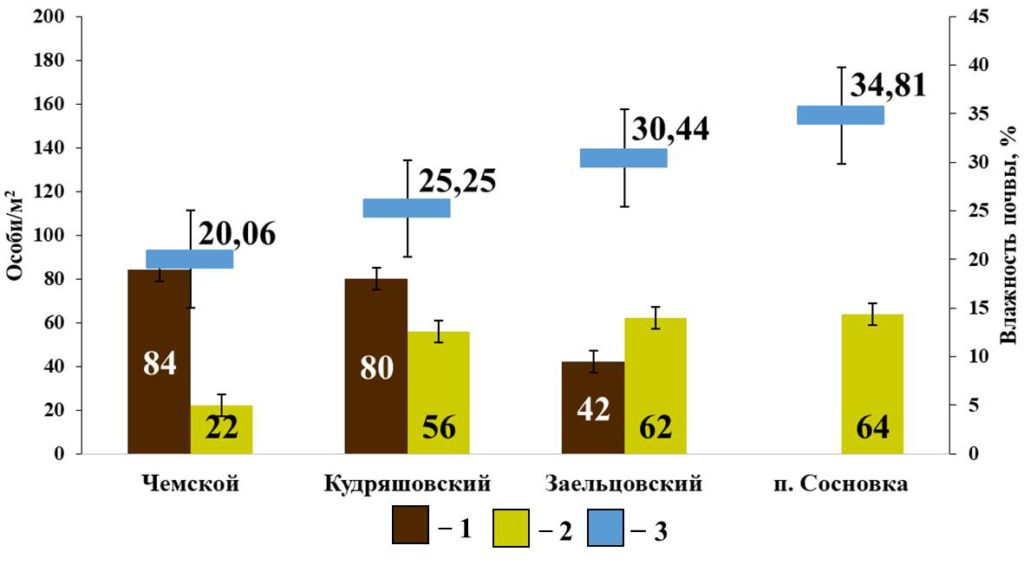
Рисунок 6. Зависимость плотности населения собственно-почвенных дождевых червей
от влажности почвы в сосновых борах
Условные обозначения: 1 — A. caliginosa caliginosa; 2 – E. nordenskioldi pallida; 3 – Влажность почвы
В мелколиственных лесах с серыми лесными почвами выявлены положительные корреляционные взаимосвязи между суммарной плотностью поверхностнообитающих червей и влажностью почвы (rs=0.89; p <0.05; n=5) (рис. 7). С другими характеристиками почвы значимые взаимосвязи не выявлены.
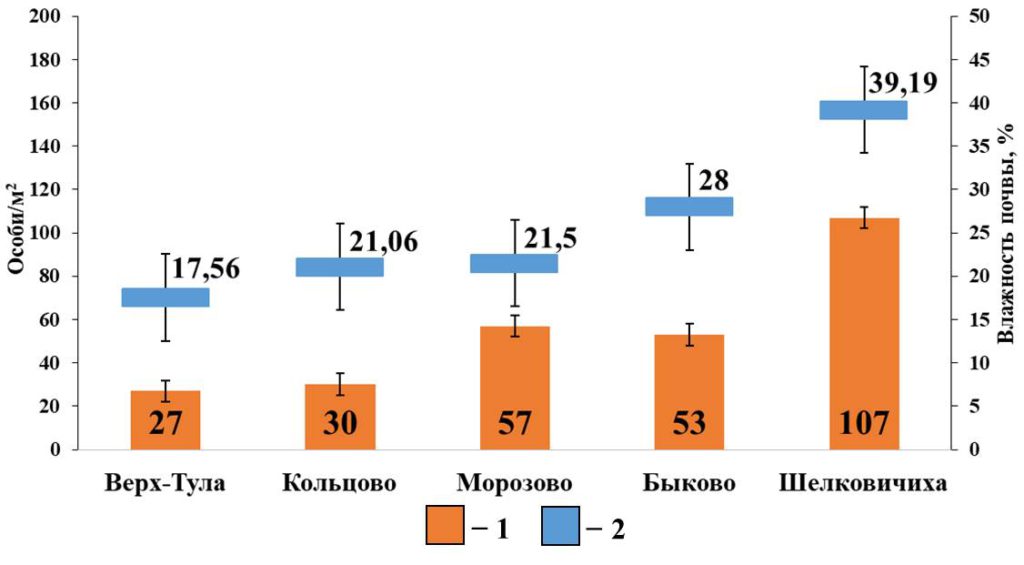
Рисунок 7. Зависимость плотности населения поверхностнообитающих дождевых червей от влажности почвы в мелколиственных лесах
Условные обозначения: 1 — Плотность населения поверхностнообитающих червей; 2 – Влажность почвы
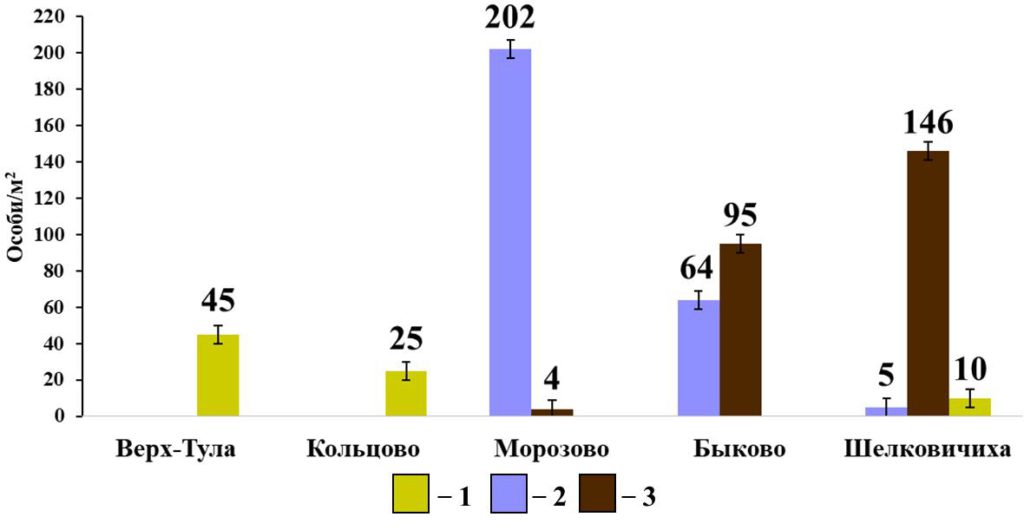
Рисунок 8. Разнообразие видового состава и плотности населения собственно-почвенных дождевых червей в мелколиственных лесах
Примечание: 1 — E. nordenskioldi pallida; 2 – O. lacteum; 2 – A. caliginosa caliginosa
В группе собственно-почвенных червей, как для суммарной плотности населения, так для отдельных видов, корреляционные взаимосвязи со свойствами почвы не установлены. Однако следует отметить, что в отличие от боров видовой состав и плотность населения этой группы дождевых червей сильно варьируют в мелколиственных лесах (рис. 8). В лесу окрестностей поселка Морозово с мощным гумусовым горизонтом (19.9 см) и слабокислой реакцией почв (pHводн.= 6.22) преобладает O. lacteum, в лесах окрестностей Быкова и Шелковичихи с кислой реакцией почв (pHводн.= 5.68; 5.42, соответственно) и менее развитым гумусовым горизонтом (12.3 см и 9.9 см, соответственно) – A. c. caliginosa. Плотность населения E. n. pallida в мелколиственных лесах заметно ниже, чем в сосновых борах и не объясняется измеренными характеристиками почвы.
Морфометрический анализ E. n. nordenskioldi. В ходе данного исследования было отмечено, что особи подвида E. n. nordenskioldi, относящегося к почвенно-подстилочным червям, заметно различаются по своим размерам: визуально возможно выделить мелкую и крупную формы (рис. 9). Результаты морфометрического анализа выборок червей разных размерных форм приведены ниже (табл. 7).

Рисунок 9. Полиморфизм E. n. nordenskioldi
Условные обозначения: 1 – крупная форма, почва; 2 – Кольцово, валежник; 3- Быково, валежник; 4 – Верх-Тула, почва.
Таблица 7. Морфометрический анализ особей E. n. nordenskioldi (X ± SE)
| № выборки | Место сбора | Число особей | Число сегментов | Длина тела, мм | Ширина тела, мм | Длина пояска, мм | Ширина пояска, мм | Вес, г |
| Мелкая размерная форма | ||||||||
| 1 | Верх-Тула, почва | 11 | 114±5 | 48.7±1.5 | 2.7±0.1 | 4.2±0.2 | 2.9±0.1 | 0.2±0.01 |
| 2 | Быково, валежник | 20 | 106±1 | 53.5±1.0 | 3.1±0.1 | 4.4±0.1 | 3.2±0.1 | 0.25±0.01 |
| 3 | Кольцово, валежник | 17 | 102±2 | 61.0±1.7 | 3.2±0.1 | 3.7±0.1 | 3.2±0.1 | 0.3±0.01 |
| Крупная размерная форма | ||||||||
| 4 | Боры | 9 | 133±7 | 114.2±5.6 | 6.0±0.3 | 6.7±0.4 | 4.9±0.2 | 1.8±0.2 |
| 5 | Мелколист. леса | 7 | 126±5 | 104.9±3.9 | 5.2±0.1 | 6.1±0.2 | 5.3±0.2 | 1.5±0.2 |
Наибольшее число признаков (5-6), по которым выявлены статистически значимые различия, было установлено при сравнении выборок крупных и мелких размерных форм E. n. nordenskioldi. При сопоставлении выборок мелких форм между собой установлено не более 3-х признаков, по которым они статистически значимо различаются. Выборки крупных форм для боров и мелколиственных лесов между собой значимо не различаются (табл. 8). Поэтому в данной работе мы приводим показатели плотности населения отдельно и для крупной и для мелкой формы E. n. nordenskioldi.
Таблица 8. Перечень статистически достоверных различий между выборками
(тест Уитни-Манна, p < 0.01)
| № выборки | 2 | 3 | 4 | 5 |
| 1 | КС, Ш, В | КС, Д, Ш, В | Д, Ш, ДП, ШП, В | Д, Ш, ДП, ШП, В |
| 2 | — | Д, ДП, В | КС, Д, Ш, ДП, ШП, В | КС, Д, Ш, ДП, ШП, В |
| 3 | — | КС, Д, Ш, ДП, ШП, В | КС, Д, Ш, ДП, ШП, В |
Примечание: условные обозначения см. раздел материалы и методы
ОБСУЖДЕНИЕ
При исследовании полиморфизма E. n. nordenskioldi следует отметить, что мелкая форма в основном населяет валежник, а в почве встречается редко, в то время как крупная форма обитает в почве, нередко на глубине более 15-20 см, хотя питается на поверхности. Возникло предположение, что крупные особи относятся к червям-норникам. Подобная точка зрения уже была предложена в период исследований лесов Западного Саяна и почв Якутии (Перель, 1994; Боескоров, 2004), где также были обнаружены крупные и мелкие формы данного подвида, причем крупная форма была обозначена как норная. Также следует отметить, что в сборах дождевых червей, сделанных Ю.Б. Бызовой в березняках окрестностей Новосибирска, крупные особи E. n. nordenskioldi определенного размера и веса считались норными червями (Бызова, 2007).

Рисунок 10. Сходство внешнего строения между крупной формой E. n. nordenskioldi (1)
и L. terrestris (2)
Эти утверждения можно подтвердить следующим наблюдением: если сравнить крупную форму E. n. nordenskioldi с типичным представителем норных червей – L. terrestris, то можно отметить сходство в их внешней морфологии (рис. 10). В частности, у крупной формы отчетливо выражено уплощение тела и ослабление пигментации в направлении от головного конца к хвостовому, в то время как у мелких червей явно выраженного уплощения тела нет, а пигментация распределена по телу весьма однородно. Однако однозначно утверждать, что крупною форму E. n. nordenskioldi, обнаруженную в Новосибирской области, можно отнести к норным червям пока невозможно, т.к. необходимы детальные исследования внутренней морфологии, а также поиск длинных вертикальных ходов в почве, типичных для норников.
Подобное исследование было проведено и для вида O. lacteum. В Новосибирской области обитают крупные и мелкие формы вида, имеющие достоверные различия не только по морфологическим признакам, но и на молекулярно-генетическом уровне (Шеховцов, Ермолов и др., 2020). Но крупная форма O. lacteum в основном приурочена к пойменным биотопам, а в лесах встречается только мелкая форма.
Население дождевых червей в сосновых борах и мелколиственных лесах в целом оказалось сходным по видовому составу и соотношению жизненных форм, но плотность червей в мелколиственных лесах значительно выше, чем в борах. Даже в сухом низкотравном березовом лесу (Верх-Тула) плотность населения была выше, чем в самом «бедном» по населению бору (Барышево). Однако население дождевых червей в высокотравно-папоротниковых борах заметно разнообразнее, чем в темнохвойных лесах (Гераськина, 2016б) и Южной тайге (Крылова и др., 2011), где, как правило, собственно-почвенные черви отсутствовали вовсе, а подстилочные и почвенно-подстилочные представлены одним видом. Березово-осиновые леса лесостепного Приобья Новосибирской области хоть и имеют разнообразный видовой состав и высокую плотность населения дождевых червей, но уступают по этим показателям мелколиственным и широколиственным лесам в Eвропейской части РФ (Перель, 1958; Аккумуляция…, 2018; Гераськина, 2016а). Главное отличие комплексов жизненных форм дождевых червей в Новосибирской области от упомянутых выше регионов – отсутствие типичных норных форм (если не считать таковой крупную форму E. n. nordenskioldi). Из них здесь крайне редко встречается L. terrestris, который населяет антропогенные территории, а в естественных местообитаниях не встречается (Ермолов, 2018а, 2019).
В исследованных лесах плотность населения собственно-почвенных червей намного превышает таковую поверхносто-обитающих. Большая часть населения подстилочных и почвенно-подстилочных червей сосредоточена в лесном валежнике, которые даже размножаются в нем, о чем свидетельствуют найденные коконы (Гераськина, 2016в; Ермолов, 2018а). По сути, валежник обеспечивает разнообразие поверхностнообитающих дождевых червей в лесных экосистемах. В разных биотопах соотношение подстилочных и почвенно-подстилочных форм дождевых червей может различаться в валежниках, но при этом видовой состав практически всегда сходен с таковым в почве (табл. 1, 2, 3, 4). Собственно-почвенных червей также можно обнаружить в валежнике. В основном они населяют сильно разложившиеся стволы, где представлены ювенильными особями, которые обитают близко к поверхности почвы. Половозрелые особи встречаются в валежниках заметно реже и, видимо, используют его как временное местообитание, например, заползают туда при выходах из почвы во время дождя (Чекановская, 1960; Гераськина, 2016г).
Также следует отметить, что в почве среди поверхностнообитающих червей плотность населения видов-космополитов (D. r. tenuis, D. octaedra, L. rubellus) и азиатского подвида (E. n. nordenskioldi) примерно одинакова в разных местообитаниях, а среди собственно-почвенных червей космополиты (O. lacteum, A. c. caliginosa) заметно превышают по численности азиатский подвид E. n. pallida. У отдельных видов подстилочных и почвенно-подстилочных червей нет приуроченности к определенным факторам почвы, а их суммарная плотность напрямую зависит от влажности почвы, так как именно эти группы обитают у самой поверхности почвы и поэтому более чувствительны к пересыханию (Чекановская, 1960; Перель, 1979; Гераськина, 2016г). При высокой влажности черви этой экологической группы обильно присутствуют даже в кислой почве, особенно в мелколиственных лесах (pHводн.= 5.42), где подстилка более питательна по сравнению с борами (Holdsworth et al., 2008). Но, тем не менее, поверхностнообитающие черви оказывают меньшее влияние на pH почвы, и поэтому зависимость их плотности населения от данного фактора требует более детальных исследований. С мощностью подстилки прямых корреляций не обнаружено, однако в лесах с присутствием L. rubellus (Заельцовский бор, Быково) отмечена минимальная толщина подстилки, так как этот вид более интенсивно перерабатывает подстилку, чем другие подстилочные и почвенно-подстилочные виды (Голованова и др., 2018). У собственно-почвенных червей в борах наблюдалась корреляция между плотностью населения отдельных видов и влажностью почвы. Обратная корреляция A. c. caliginosa с влажностью почвы, вероятно, объясняется тем, что подвид способен к летней диапаузе и поэтому может лучше переносить засуху, чем другие виды дождевых червей (Перель, 1975, 1979). Но в более влажных почвах увеличивается плотность населения других видов, которые могут превосходить A. c. caliginosa по численности. В мелколиственных лесах значимых корреляций отмечено не было. При этом изученные мелколиственные леса сильно различались по видовому составу собственно-почвенных червей: в березово-осиновом лесу в окрестностях поселка Морозово преобладал O. lacteum, что можно объяснить мощным гумусовым горизонтом и слабой кислотностью почвы. Обычно этот вид преобладает в переувлажненных почвах, но как сказано выше, в мелколиственных лесах исследованного региона, почва которых недостаточно увлажнена, особи O. lacteum представлены только мелкой размерной формой, а более влажные биотопы (поймы рек) населяет крупная размерная форма вида (Шеховцов, Ермолов и др., 2020). Также было установлено, что размер взрослых особей O. lacteum коррелирует с влажностью почвы (Ермолов, неопубл. данные), а мощный гумусовый горизонт обеспечивает присутствие вида, так как является для него основным местообитанием и источником пищи (Чекановская, 1960; Перель, 1979). В более кислых, но также и более влажных местообитаниях встречается A. c. caliginosa, в сухих лесах со слабокислой почвой обнаружен только подвид E. n. pallida, который, видимо, способен выживать в почвах с небольшим содержанием влаги и маломощным гумусовым горизонтом.
ЗАКЛЮЧЕНИЕ
В сосновых борах и мелколиственных лесах лесостепного Приобья Новосибирской области видовой состав дождевых червей во многом сходен, но суммарная плотность населения червей заметно выше в мелколиственных лесах. В большинстве местообитаний значительную долю населения составляют собственно-почвенные среднеярусные формы, лишь в одном случае (мелколиственный лес в п. Морозово), доминантами оказались собственно-почвенные верхнеярусные черви. Также в некоторых лесах высока доля поверхностнообитающих дождевых форм червей.
Исследованные в лесах валежники классифицированы на три группы, в зависимости от преобладания определенных жизненных форм дождевых червей. Больше всего обнаружено валежников с преобладанием подстилочных форм, меньше всего с преобладанием собственно-почвенных форм. Но при одинаковом соотношении жизненных форм валежники в пределах одной группы могут значительно различаться по видовому составу червей.
В ходе исследования подтверждено, что в лесных экосистемах наиболее значимым фактором для дождевых червей является влажность почвы, особенно для поверхностнообитающих жизненных форм. Для собственно-почвенных форм дождевых червей также важна мощность гумусового горизонта.
После проведения морфометрического анализа по внешним морфологическим признакам обосновано выделение крупной и мелкой размерных форм подвида Eisenia n. nordenskioldi. Мелкие формы данного подвида являются типичным почвенно-подстилочными червями, населяющими почву и валежник. Крупная форма, возможно, относится к червям-норникам.
Проведенное исследование предоставляет начальные сведения о населении дождевых червей в лесных экосистемах лесостепного Приобья Новосибирской области. В дальнейших работах планируется провести более детальные исследования лесов каждого типа, уделяя особое внимание сосновым борам и смешанным лесам, так как именно в них обитают виды дождевых червей, имеющие ограниченный ареал и приуроченные исключительно к азиатской части России.
БЛАГОДАРНОСТИ
Автор выражает искреннюю признательность и благодарность аспиранту Кафедры общей биологии и экологии НГУ Ким-Кашменской Марии Никитичне за многочисленные консультации и поддержку на основных этапах работы; профессору Сергееву Михаилу Георгиевичу и старшему преподавателю Молодцову Владимиру Владимировичу (НГУ, Кафедра общей биологии и экологии) за наставничество, взаимо-понимание, помощь в организации исследования, технические и научные советы по обработке данных и консультирование по организационным вопросам, а также рецензентам за ценные советы и справедливые замечания.
СПИСОК ЛИТЕРАТУРЫ
Аккумуляция углерода в лесных почвах и сукцессионный статус лесов // под ред. Н. В. Лукиной. М.: Товарищество научных изданий КМК, 2018. 232 с.
Атлас Новосибирской области // под ред. В.М. Кравцова. М.: Роскартография, 2002. 56 с.
Боескоров В.С. Экологические условия обитания дождевого червя Eisenia nordenskioldi, Eisen в мерзлотных почвах Якутии // Автореф. … дисс. канд. биол. наук. Улан-Удэ: ИОиЭБ СО РАН, 2004. 24 с.
Бызова Ю.Б. Дыхание почвенных беспозвоночных. М.: Товарищество научных изданий КМК, 2007. 328 с.
Воробьева Л.А., Ладонин Д.В., Лопухина О.В., Рудакова Т.А., Кирюшин А.В. Химический анализ почв. Вопросы и ответы. М., 2012. 186 с.
Всеволодова-Перель Т.С. Дождевые черви фауны России. Кадастр и определитель. М.: Наука, 1997. 102 с.
Вэн Райзин Дж. Классификация и кластер. М.: Мир, 1980. 390 с.
Ганин Г.Н. Структурно‑функциональная организация сообществ мезопедобионтов юга Дальнего Востока России. Владивосток: Дальнаука, 2013. 380 с.
Гераськина А.П. Дождевые черви (Oligochaeta, Lumbricidae) окрестностей пос. Домбай Тебердинского заповедника (Северо-западный Кавказ, Карачаево-Черкессия) // Труды Зоологического института РАН. 2016а. Т. 320. № 4. С. 450-466.
Гераськина А.П. Население дождевых червей (Lumbricidae) в основных типах темнохвойных лесов Печеро-Илычского заповедника // Зоологический журнал. 2016б. Т. 95. № 4. С. 394-405.
Гераськина А.П. Проблемы количественной оценки и учета фаунистического разнообразия дождевых червей в лесных сообществах // Russian Journal of Ecosystem Ecology. 2016в. Vol. 2. № 2. P. 1-9.
Гераськина А.П. Экологическая оценка динамики комплекса дождевых червей (Lumbricidae) в ходе восстановительных сукцессий: монография. Смоленск: Изд. СГМУ, 2016г. 149 с.
Голованова Е.В., Князев С.Ю., Карабан К. Есть ли преимущества у аборигенного вида дождевых червей по сравнению с видами вселенцами в Западной Сибири? // Материалы XVIII Всероссийского совещание по почвенной зоологии: Тез. докл. 22–26 октября 2018 г. Москва, 2018. С. 60-61.
ГОСТ 28268-89. Почвы. Методы определения влажности, максимальной гигроскопической влажности и влажности устойчивого завядания растений. 1990. М.: Стандартинформ, 2006. 14 с.
Ермолов С.А. Особенности распределения жизненных форм дождевых червей (Lumbricidae) лесостепного Приобья // Научные основы устойчивого управления лесами: Материалы Всероссийской научной конференции. Москва: ЦЭПЛ РАН, 2018а. С. 43-45.
Ермолов С.А. Фаунистическое разнообразие и экология дождевых червей в биотопах речных долин лесостепного Приобья // 56-я Международная научная студенческая конференция МНСК-2018: Тез. докл. 22–27 апреля 2016 г. Новосибирск, 2018б. C. 155.
Ермолов С.А. Биотопическое распределение дождевых червей (Oligochaeta, Lumbricidae) в малых речных долинах лесостепного Приобья // Russian Journal of Ecosystem Ecology. 2019. Vol. 4. № 2. С. 1-18.
Ким-Кашменская М.Н. Фауна дождевых червей (Oligichaeta, Lumbricidae) долина р. Бердь в Присалаирье // Биоразнообразие: глобальные и региональные процессы: Тез. докл. Всерос. конф. молодых ученых с междунар. участием. 23–27 июня 2016 г. Улан-Удэ, 2016. С. 242-243.
Крылова Л.П., Акулова Л.И., Долгин М.М. Дождевые черви (Oligochaeta, Lumbricidae) Таежной зоны республики Коми. Сыктывкар, 2011.104 с.
Методы почвенно-зоологических исследований // под ред. М.С. Гилярова. М.: Наука, 1975. 281 с.
Мугако А.Л. Природа Новосибирской области. Новосибирск: Новосибирский государственный краеведческий музей, 2008. 40 с.
Мусиенко И.Е. Оценка видового разнообразия семейства Lumbricidae в Таштагольском районе Горной Шории // 57-ая Международная научная студенческая конференция МНСК-2019: Тез. докл. 14–19 апреля 2019 г. Новосибирск, 2019. С. 25.
Пенев Л.Д., Васильев А.И., Головач С.И., Квавадзе Е.Ш. Вдовой состав и классификация группировок дождевых червей (Oligochaeta, Lumbricidae) дубрав русской равнины // Зоологический журнал. 1994. Т. 73. № 2. С. 23-37.
Перель Т.С. Зависимость численности и видового состава дождевых червей от породного состава лесонасаждений // Зоологический журнал. 1958. Т. 37. № 9. С. 1307-1314.
Перель Т.С. Жизненные формы дождевых червей (Lumbricidae) // Журнал общей биологии. 1975. Т. 36. № 2. С. 189-202
Перель Т.С. Распространение и закономерности распределения дождевых червей фауны СССР. М., Наука, 1979. 272 с.
Перель Т.С. Дождевые черви (Oligochaeta, Lumbricidae) в лесах Западного Саяна (с описанием нового вида) // Зоологический журнал. 1994. Т. 73. № 2. С. 18-22.
Почвенная карта Новосибирской области // под ред. К.С. Байкова. Новосибирск. Изд. ИПА СО РАН, 2007.
Рапопорт И.Б. Сезонная активность дождевых червей (Oligochaeta, Lumbricidae) пояса широколиственных лесов кабардино-балкарского государственного высокогорного заповедника и прилегающих территорий (Центральный Кавказ) // Известия самарского научного центра Российской академии наук. 2010. Т. 12. № 1(5). С. 1345-1348.
Чекановская О.В. Дождевые черви и почвообразование. М.; Л., Изд-во АН СССР 1960. 208 с.
Шашков М.П. Фауна дождевых червей (Lumbricidae) заповедника «Калужские засеки» // Труды государственного природного заповедника «Калужские засеки». Вып. 1. 2003. С. 90-93.
Шеховцов С.В., Базарова Н.Э., Берман Д.И., Булахова Н.А., Голованова Е.В., Коняев С.В., Кругова Т.М., Любечанский И.И., Пельтек С.Е. ДНК-штрихкодирование: сколько видов дождевых червей живет на юге Западной Сибири? // Вавиловский журнал генетики и селекции. 2016. Т. 20. № 1. С. 125-130.
Шеховцов С.В., Ермолов С.А., Держинский Е.А., Полубоярова Т.В., Ларичева М.С., Пельтек С.Е. Генетическая и размерная изменчивость Octolasion tyrtaeum (Lumbricidae, Annelida) // Письма в Вавиловский журнал генетики и селекции. 2020. Т. 6. № 1. С. 5-9.
Holdsworth A.R., Frelich L.E., Reich P.B. Litter decomposition in earthworm-invaded northern hardwood forests: Role of invasion degree and litter chemistry // Ecoscience. 2008. Vol. 15. No. 4. P. 536-544.
Ashwood F., Vanguelova E.I., Benham S., Butt K.R. Developing a systematic sampling method for earthworms in and around deadwood // Forest Ecosystems. 2019. Vol. 6. No. 33. P. 1-12.
Рецензент: к.б.н., с.н.с. Гераськина А.П.





