- DOI 10.31509/2658-607x-202143-88
- УДК 630*181.351, 574.472
Мозаичность фитоценозов хвойно-широколиственных лесов Валуевского лесопарка
![]()
Е. В. Тихонова1, Г. Н. Тихонов2
1Центр по проблемам экологии и продуктивности лесов Российской академии наук Россия, 117997 Москва, ул. Профсоюзная, 84/32, стр. 14
2University of Helsinki, Faculty of Biological and Environmental Sciences, Organismal and Evolutionary Biology Research Programme, PL 65 (Viikinkaari 1), 00014 Finland
E-mail: tikhonova.cepl@gmail.com
Поступила в редакцию: 15.01.2021
После рецензирования: 17.03.2021
Принята к печати: 18.08.2021
Актуальность и цель. Отечественная геоботаника имеет глубокие традиции изучения неоднородности растительного покрова. Однако число исследований мозаичности сложно организованных хвойно-широколиственных лесов весьма ограничено и полученные результаты неоднозначны. Кроме этого, назрела необходимость разобраться с «терминологическим вопросом». Целью настоящего исследования была оценка мозаичности растительного покрова широко распространенных на Москворецко-Окской равнине типов лесных фитоценозов.
Материал и методы. Приведен обзор публикаций по внутриценотической неоднородности лесного покрова и сделана попытка систематизировать имеющуюся терминологию. Полевые исследования проведены на территории Валуевского лесопарка (Новомосковский АО, г. Москва). В работе использованы традиционные геоботанические методы и различные современные методы статистического анализа данных.
Результаты и заключение. Полученные результаты показывают, что различия в растительности намного сильнее выражены на уровне фитоценозов, чем на уровне микрогруппировок. Подтверждено наиболее сильное эдификаторное воздействие ели. Возможными причинами, определяющими слабую дифференциацию растительности на уровне микрогруппировок, являются: влияние соседних микрогруппировок, недостаточная для заметной трансформации растительности продолжительность воздействия отдельных деревьев и наследование в структуре и видовом составе предшествующих состояний за длительный период времени.
Ключевые слова: хвойно-широколиственные леса, мозаичность, фитоценоз, микрогруппировка, Москворецко-Окская равнина
Современные представления о многоуровневой (иерархической) организации растительного покрова позволяют рассматривать его как совокупность разномасштабных пространственных (или территориальных) единиц (Сочава, 1972; Маслов, 1990; Заугольнова, 1999). Изучение неоднородности растительного покрова — его фундаментального свойства — имеет глубокие традиции в отечественной фитоценологии (Корчагин, 1976). Признавая фитоценоз основной единицей растительного покрова, исследователи выделяли и описывали его структурные элементы и анализировали причины, вызывающие неоднородность. А. П. Шенников в 1920-х годах ввел в научный обиход термины «мозаичность» и «микрогруппировка». Учение о мозаичности растительного покрова получило свое развитие в работах П. Д. Ярошенко (1953, 1958, 1969), позже А. А. Корчагин (1976) в очень подробном обзоре обобщил результаты исследований внутриценотической неоднородности. Следует отметить, что ученые уже давно обращали внимание на проблему терминологии, на разное понимание объема и содержания используемых понятий, когда одни и те же пространственные единицы назывались по-разному или же один термин использовался для обозначения разных объектов. На основе обзора результатов исследований, связанных с изучением внутриценотической неоднородности лесных сообществ, мы попробуем систематизировать терминологию, для чего сопоставим определения ключевых понятий и подчеркнем расхождения в понимании терминов разными авторами.
П. Д. Ярошенко (1953, с. 6.) определил фитоценоз (растительную группировку, растительное сообщество), как «совокупность растений, занимающих определенный участок и объединенных взаимодействиями со средой, а через посредство среды и между собою». Он особо подчеркивал, что сообщество должно рассматриваться не как простая сумма особей, а как целое, обладающее своими новыми специфическими качествами, что в настоящее время определяется понятием эмерджентности. Под микрогруппировками (или микрофитоценозами) П. Д. Ярошенко (1969, с. 45) понимал «мелкие сочетания растений внутри сообщества», которые включают все ярусы сообщества. Структурные элементы фитоценоза, тождественные микрогруппировкам, М. И. Сахаров (1950) называл «ценоэлементами», однако П. Д. Ярошенко (1953, с. 45) считал, что «термин “микрогруппировка” излишне заменять каким-либо другим».
А. А. Корчагин для пространственной единицы, совпадающей по объему и содержанию с микрогруппировкой Ярошенко, предложил термин микроценоз, которому он дает следующее определение (Корчагин, 1976, с. 110). Микроценоз — это закономерно сложившаяся, наименьшая по размерам структурная единица горизонтального расчленения всего сообщества, включающая все ярусы, обособленная по всей вертикальной толще во всех ярусах от других окружающих ее микроценозов этого же сообщества и отличающаяся от них по составу, экобиологическим свойствам своих компонентов, фитосреде, структуре, динамике, а также находящаяся во взаимосвязи и взаимодействии (обмене веществом и энергией) с условиями среды и с другими микроценозами и обладающая некоторой целостностью.
Под микрогруппировкой же А. А. Корчагин (1976, С. 114) понимал структурные элементы, которые можно выделить в пределах каждого яруса, т. е. относящиеся к внутриярусной мозаике. Отметим, что Л. О. Карпачевский с соавторами (2007) предлагали использовать термин «предельные элементы» для обозначения такого рода объектов (ветровально-почвенных комплексов, муравьиных куч, валежа).
Н. В. Дылис ввел понятие парцеллы как элемента горизонтальной неоднородности биогеоценоза. «Биогеоценотическими парцеллами называются структурные части горизонтального расчленения биогеоценоза, отличающиеся друг от друга составом, структурой и свойствами своих компонентов, спецификой их связей и материально-энергетического обмена. Обособлены парцеллы друг от друга в пространстве на всю вертикальную толщу биогеоценоза» (Дылис, 1969, с. 12). Он указывает достаточно большой диапазон размеров парцелл — от нескольких до десятков и сотен квадратных метров. Из данного уточнения следует, что Н. В. Дылис не отождествлял парцеллу исключительно с отдельным деревом и его фитогенным полем, но также с группой деревьев (куртиной) или участком ветровального окна. По мнению Н. В. Дылиса, одиночные деревья могут формировать парцеллу при условии их хорошего развития и существенных отличий от своего окружения.
Н. В. Дылис отмечал, что парцеллы по объему и содержанию близки к ценоэлементам Сахарова, а растительный компонент парцеллы соответствует понятию микрогруппировки Ярошенко (или микроценоза Корчагина). По оценкам Н. В. Дылиса (1969), лесные биогеоценозы подзоны хвойно-широколиственных лесов могут включать от 2 до 14 парцелл. Наименьшее число парцелл свойственно лесам с бедным составом древесного яруса и слабо развитым ярусом кустарников. Простую парцеллярную структуру имеют молодые лиственные леса с однородной структурой древостоя и молодые сомкнутые ельники. Большее число парцелл (5–8) характерно для позднесукцессионных широколиственно-еловых лесов, а также для производных березняков и осинников на стадиях их смены елью. Однако наиболее сложная парцеллярная структура отмечена в средневозрастных и приспевающих широколиственно-еловых лесах, в которых при регулярном проведении санитарных и проходных рубок создаются производные парцеллы.
Почвенный компонент парцелл был детально изучен Л. О. Карпачевским (Карпачевский, 1977; Карпачевский и др., 1980). Л. О. Карпачевский (1977) указывал на некоторую нечеткость определения парцеллы, данного Н. В. Дылисом, и в качестве «рабочей» предложил следующую формулировку: «парцелла — структурная часть БГЦ, площадь которой определяется границами распространения данных доминантов и данных (конкретных, индивидуальных) эдификаторов» (Карпачевский, 1977, с. 46). Эдификатором (центром, «организатором») «может быть дерево, куст, даже геометрический центр окна» (Карпачевский и др., 1980, с. 3), вокруг этого центра формируется фитогенное поле (в понимании Уранова, 1965). Таким образом, парцеллу Л. О. Карпачевский отождествляет с фитогенным (биогеоценотическим) полем, а ее почвенный компонент — с тессерой Г. Иенни (Карпачевский, 1977; Карпачевский и др., 1980). Каждое растение обладает фитогенным полем, и в биогеоценозах поля разного порядка находятся во взаимодействии друг с другом. Однако эдификатор парцеллы формирует наиболее сильное биогеоценотическое поле, приводящее к закономерному изменению свойств почв по радиусу парцеллы (Карпачевский и др., 1980). Парцеллы было предложено называть по эдификатору и доминантам напочвенного покрова: елово-волосистоосоковая, березово-мертвопокровная, крупнопапоротниковая в окне и т. д.
Достаточно единодушны разные авторы в понимании взаимоотношений между парцеллами (микрогруппировками, ценоэлементами) и биогеоценозом (фитоценозом). Л. О. Карпачевский с соавторами (1980) отмечали, что парцеллы представляют собой подсистемы в более крупной системе — биогеоценозе, элементом которой они являются. Примерно о том же говорил А. А. Корчагин (1976, С. 12): «Ценоэлементы не представляют самостоятельных единиц, а являются лишь частями, кирпичиками растительного сообщества — пространственно, экологически и фитоценотически единой, до некоторой степени целостной системы, первичной конкретной единицы растительного покрова». Н. В. Дылис (1969) отмечает, что существование каждой парцеллы зависит от окружающих ее парцелл, входящих в данный биогеоценоз. Любое изменение окружения приводит к нарушению связей между компонентами парцеллы, что в конечном итоге влечет за собой глубокую перестройку и смену парцеллы на новую структуру. На этом основании Н. В. Дылис заключает, что «представление о биогеоценотических парцеллах как специфических и по природным свойствам достаточно сильно дифференцированных ячейках биогеоценозов не противоречит представлению об известной целостности биогеоценоза как элементарного составляющего биогеосферы, а лишь отражает природные закономерности его горизонтального сложения, его мозаичность» (Дылис, 1969, с. 36).
Большинство проведенных исследований внутриценотической неоднородности растительного покрова связаны с анализом фитогенного поля деревьев-эдификаторов. А. А. Уранов (1965), изучая взаимодействия между растениями, пришел к выводу, что воздействие одного растения на другие совершается в основном путем изменения условий обитания («среды жизни»). Он сформулировал понятие фитогенного поля растения как «части пространства, в пределах которой среда приобретает новые свойства, определяемые присутствием в ней данной особи растения» (Уранов, 1965, с. 251) и обозначил два основных вопроса при изучении фитогенного поля: 1) как изменяет каждое растение среду; 2) как реагируют на эти изменения другие члены растительного сообщества.
Надо отметить, что почти одновременно с теорией А. А. Уранова о фитогенном поле была разработана очень близкая к ней концепция, получившая название «single-tree influence circles» (Zinke, 1962). В ней описываются зоны влияния отдельных деревьев, представляющие радиально-симметричные структуры, в пределах которых закономерно меняются свойства почв. Эта концепция получила широкую известность за рубежом, но в публикациях отечественных ученых она практически не упоминается.
С момента создания А. А. Урановым теории фитогенного поля шло активное накопление фактических данных по разным видам растений из разных типов растительных сообществ и из разных географических регионов, и одновременно разрабатывались методы количественной оценки параметров фитогенного поля растений. Надо отметить, что преобладающее число исследований посвящено фитогенным полям деревьев.
В структуре фитогенных полей деревьев обычно выделяют несколько зон: пристволовую, внутреннюю (основную), ограниченную проекцией кроны, и внешнюю, выходящую за пределы проекции кроны (Маслов, 1986; Черняева, Викторов, 2016). Некоторые авторы выделяют как отдельную зону краевую полосу проекции кроны (Крышень, 1998). В лесных фитоценозах фитогенные поля отдельных деревьев накладываются друг на друга с образованием зон интерференции (Маслов, 1986; Ястребов, Лычная, 1993; Черняева, Викторов, 2016).
Понятие «напряженность фитогенного поля» отражает величину изменений, вносимых особью растения в разных точках фитогенного поля, снижаясь по мере удаления от этой особи (Ипатов, Кирикова, 2001).
Были определены основные экологические и ценотические параметры, на которые оказывает влияние фитогенное поле деревьев: освещенность, температура воздуха и почвы, количество и химический состав осадков, влажность, богатство и кислотность почвы, характер опада и др. (Økland et al., 1999; Ипатов, Кирикова, 2001; Лебедева и др., 2007). Эти факторы в пределах фитогенного поля закономерно и сильно изменяются и могут характеризовать его напряженность. Напряженность фитогенного поля можно также оценить, используя в качестве индикатора растительность нижних ярусов (Ашик, Тиходеева, 2006).
Сила влияния дерева (т. е. глубина трансформации среды) обратно пропорциональна расстоянию от дерева и прямо пропорциональна размеру и возрасту дерева (Котов, 1983; Самойлов, Тархова, 1993; Крышень, 1998; Ашик, Тиходеева, 2006).
Среди видов растений наиболее сильным фитогенным полем обладают деревья, и в европейской части России в этом плане обычно выделяют ель (Picea sp.). В. С. Ипатов и Л. А. Кирикова (2001, с. 102) отмечают, что «в зоне максимальной напряженности своего фитогенного поля ель в отличие от многих других древесных пород не изменяет напочвенный покров, а уничтожает его; в результате при высокой сомкнутости древостоя формируются мертвопокровные ельники». Однако фитогенное поле закономерно изменяется с возрастом дерева. В исследовании фитогенного поля ели в сосняках зеленомошных Неруссо-Деснянского полесья было показано, что в онтогенезе деревьев происходит значительное увеличение размеров фитогенных полей (Киричок и др., 2008). С возрастом ели также увеличивается высота прикрепления кроны, и с этим параметром положительно связаны проективное покрытие и видовое богатство травяно-кустарничкового яруса подкроновых участков. На связь высоты прикрепления кроны ели со степенью развития напочвенного покрова обращали внимание и другие авторы (Ипатов, 2007; Лебедева и др., 2007).
Минимальные величины освещенности (7–10% от освещенности на фоновых участках) отмечаются в пристволовой части фитогенного поля ели. Увеличение освещенности происходит медленно до середины кроны, а после резко ускоряется (Ипатов, Кирикова, 2001). Такой характер изменения освещенности в пределах фитогенного поля связан с конусовидной формой кроны ели, когда протяженность кроны максимальна у ствола дерева и уменьшается в направлении периферии кроны (Ипатов и др., 2009).
Было показано (Ипатов, Кирикова, 2001), что задержание осадков кронами елей зависит от интенсивности дождя. Если в моросящий дождь кроны крупных елей задерживают до 95% осадков, то в сильный дождь — до 70% осадков. Меньше всего осадков при моросящем дожде поступает в пристволовую часть — около 3% от выпавших за пределами кроны. При сильном дожде заметно увеличивается пропускание осадков в средней части кроны и у края оно достигает 60–70% выпавших на открытом месте. Стволовой сток для ели составляет в среднем всего 3% от осадков на вырубке, в то время как у осины и березы он достигает 25 и 35% соответственно (Ипатов, Кирикова, 2001). По сравнению с кронами ели, кроны сосен (Pinus sylvestris L.) пропускают намного больше осадков. Перехват осадков древесным пологом в сосняках разной полноты в среднем составляет от 16 до 32% (Петров, 1983, цит. по Маслов, 1986). Пристволовые круги сосен получают осадков на 10–30% меньше, чем межкроновые пространства (Волокитина, 1979, цит. по Маслов, 1986).
Ель оказывает заметное влияние на температурный режим в подпологовом пространстве, причем максимальные температуры под кронами елей ниже, а минимальные — не отличаются или несколько выше, чем за пределами кроны. Соответственно, различаются и перепады температур: в подкроновом пространстве разница между максимальными и минимальными температурами составляет в среднем 10 °C, в то время как на открытом месте достигает 15 °C (Ипатов, Кирикова, 2001).
Фитогенные поля липы мелколистной (Tilia cordata Mill.) были исследованы на северном пределе ее произрастания — в средней тайге Карелии (Платонова и др., 2006) и в подзоне южной лесостепи — в Белгородской области (Ашик, Тиходеева, 2006). В обеих публикациях показано влияние липы на поддержание высокого видового разнообразия посредством изменения эдафической среды ее фитогенным полем.
На юге Карелии благодаря влиянию липы обеспечивается поддержание видов бореально-неморальной и неморальной эколого-ценотических групп (ЭЦГ) (Платонова и др., 2006). Результаты анализов показали, что кислотность почвы во внутренней части фитогенного поля липы (4.9–6.0) значимо выше, чем во внешней части ее фитогенного поля (4.3–5.3) и под кронами хвойных деревьев (3.8–5.2). С помощью корреляционного анализа была выявлена разная реакция видов травяно-кустарничкового покрова в зоне фитогенного поля липы. Проективное покрытие и встречаемость Convallaria majalis L., Viola mirabilis L. и Lathyrus vernus (L.) Bernh. снижались при удалении от ствола липы, а у Vaccinium myrtillus L., Vaccinium vitis-idaea L. и Linnaea borealis L. эти показатели повышались.
В работе Е. В. Ашик и М. Ю. Тиходеевой (2006) изучались фитогенные поля липы мелколистной разных возрастных групп, произрастающих в посадках сосны обыкновенной. Сквозистость крон взрослых лип в среднем составляла 5%, а сквозистость крон сосен на фоновых участках — около 50%. Сильная затененность под пологом лип определяла слабое развитие травяного покрова, проективное покрытие которого не превышало 10%. Авторы полагают, что накопление опада под кронами лип можно объяснить снижением микробиологической активности верхнего горизонта почвы, определяющей скорость минерализации растительных остатков. Было оценено влияние фитогенного поля липы на характер распределения видов напочвенного покрова. Виды дубравной группы (Stellaria holostea L., Veronica chamaedrys L.) проявляют тяготение к кронам липы, но максимального проективного покрытия достигают на краю кроны, где лучше условия освещения и влажности и нет прямого воздействия солнечных лучей. Виды сосновой группы (Festuca ovina L., Veronica incana L., Hieracium pilosella L. и др.) и зеленые мхи ведут себя противоположным образом, они отсутствуют под кронами взрослых лип и более обильны на фоновых участках.
Фитогенные поля березы повислой (Betula pendula Roth) были исследованы в Новгородской и Тверской областях (Тарасова и др., 2003). Авторы отмечают, что сходство освещенности и параметров микроклимата под кронами лиственных деревьев делает особенно интересным анализ почв, поскольку видоспецифичное воздействие фитогенных полей может проявиться именно в изменении характеристик почвы. Результаты химических анализов почвы показали заметное влияние березы на pH и содержание в почве различных форм кальция, калия, фосфора, азота и углерода. Однако было установлено, что лишь немногие виды растений реагируют на изменения почвенных характеристик, создаваемые березой. Ю. В. Тарасова с соавт. (2003) предлагают два объяснения этого результата: небольшую величину изменений и низкую чувствительность к изменениям большинства видов растений, обладающих широкими экологическими амплитудами.
Число научных работ, в которых проводилось сравнение фитогенных полей разных видов деревьев в сходных экотопических условиях, весьма ограничено. В. Х. Лебедева с соавторами (2007) изучали фитогенные поля сосен, елей и берез в березняках черничных в Ленинградской и Псковской областях. Они показали, что средопреобразующее влияние крон елей связано со снижением освещенности и температуры, уменьшением количества проникающих осадков, образованием большого количества опада. В отличие от елей, березы оказывают слабое затеняющее действие, формируют опад хорошего качества и способствуют повышению плодородия почвы благодаря усилению деятельности микрофлоры и почвенной фауны.
И. В. Припутина с соавторами (2020) исследовали пространственную неоднородность почвенного покрова в многовидовых хвойно-широколиственных лесах на территории Приокско-Террасного заповедника (ПТЗ) на юге Московской области. В работе сравнивались виды деревьев, формирующие первый ярус: береза, сосна и ель. Полученные результаты показали, что влияние разных видов деревьев на содержание углерода (Сорг) и азота (Nобщ) наиболее заметно прослеживается в горизонте подстилки. Значимые различия получены между березой и сосной для мощности подстилки и содержания в ней Сорг. Авторы отмечают, что, исходя из имеющихся в литературе данных, они ожидали получить для дерново-подбуров ПТЗ более явные связи между содержанием Сорг и Nобщ в почвенных горизонтах и пространственно-видовой структурой древесного полога. Для исследованного биогеоценоза с нерегулярным размещением деревьев характерно формирование поверхностного опада относительно сходного состава (смешанного из хвои и листвы многих видов деревьев). Этому может способствовать «эффект воздушного переноса опадающей листвы и хвои», который снижает пространственную контрастность показателей содержания Сорг и Nобщ в горизонте подстилки. Одним из важных результатов исследования был вывод о том, что запасы углерода и азота в поверхностных горизонтах почв межкроновых участков в среднем ниже, чем под кронами деревьев.
Из обзора публикаций по внутриценотической неоднородности лесного покрова можно сделать следующие выводы: 1) большинство исследований посвящено оценке фитогенного поля одного древесного вида; 2) чаще исследовались фитогенные поля одиночных деревьев, произрастающих в контрастном окружении (на лугах, в лесах с сильно отличающимся видовым составом); 3) в большинстве исследований проводится оценка внутренней структуры фитогенного поля (с чем связан типичный дизайн эксперимента — описание растительности на площадках маленького размера, заложенных по трансекте от ствола к периферии кроны).
Цель нашего исследования — оценка мозаичности растительного покрова на уровне микрогруппировок в нескольких типах лесных фитоценозов, широко распространенных на Москворецко-Окской равнине. Нашей задачей было выявить различия между микрогруппировками, связанные с воздействием фитогенных полей разных видов-эдификаторов, как в пределах одного типа фитоценоза, так и в разных типах фитоценозов.
МАТЕРИАЛ И МЕТОДЫ
Наземные исследования проведены в пределах Валуевского лесопарка, созданного в 1935 году как часть Лесопаркового защитного пояса г. Москвы. Общая площадь исследуемой территории составляет 1156.33 га (Публичная…, 2020). Лесопарк расположен на Москворецко-Окской равнине и относится к Апрелевско-Кунцевскому ландшафту (Ландшафты …, 1997), в котором доминируют местности моренных равнин с абсолютными высотами 190–210 м. Среднегодовое количество осадков — 706 мм, среднегодовая температура 5.3 °С (Справочная информация…, 2021). В почвенном покрове преобладают дерново-подзолистые почвы на покровном суглинке. Почвообразующая порода характеризуется среднесуглинистым гранулометрическим составом (Аккумуляция…, 2018).
Характер растительности Валуевского лесопарка определяется его положением в полосе контакта двух крупных геоботанических районов — хвойно-широколиственных и широколиственных лесов (Петров, Кузенкова, 1968; Леса Южного Подмосковья, 1985), коренные типы леса на водоразделах характеризуются смешанным древостоем из липы и ели (Курнаев, 1982). Современное разнообразие лесов во многом определяется историей природопользования — границы между разными по составу древостоя лесными участками частично совпадают с границами землевладений XVIII–XIX вв. (Юрков и др., 2019), их можно проследить по сохранившимся межевым канавам. Сообщества с доминированием хвойных пород (преимущественно ели) занимают 34%, липняки — 20%, мелколиственные леса — 46% территории Валуевского лесопарка (Мучник и др., 2020). Преобладающий возраст древостоев находится в диапазоне от 60 до 110 лет (Материалы лесоустройства…, 2005). Подробная характеристика лесных сообществ с полными геоботаническими описаниями приведена в более ранней публикации (Аккумуляция…, 2018).
Наличие сильно различающихся типов фитоценозов с участием разных древесных эдификаторов в пределах однородной по экотопическим условиям территории делает Валуевский лесопарк удобным модельным объектом для исследования мозаичности растительного покрова.
Геоботанические исследования и оценка биометрических параметров деревьев проводились на квадратных площадках двух размеров: 1) для описания фитоценозов — 20×20 м; 2) для описания микрогруппировок — 5×5 м. Площадки 20×20 м закладывались в пределах всего лесного массива для определения ценотического разнообразия территории и получения характеристик типов фитоценозов.
Площадки 5×5 м располагались в пределах или рядом с некоторыми площадками 20×20 м, они закладывались вокруг дерева — центра (эдификатора) микрогруппировки и обозначались по виду дерева — Е (Picea abies), Д (Quercus robur), Б (Betula pendula), Лп (Tilia cordata). В качестве центральных деревьев выбирались деревья из первого яруса, с хорошо развитой кроной, генеративного онтогенетического состояния (g2–g3). Всего было выполнено 63 описания на площадках 20×20 м и 41 описание на площадках 5×5 м, в том числе Е — 10 (на 3-х площадках 20×20 м), Д — 10 (на 2-х площадках), Б — 9 (на 2-х площадках), Лп — 12 (на 4-х площадках). Размер площадок для описания микрогруппировок (5×5 м) был достаточным для выявления видового состава (площадь проекции кроны дерева варьировала от 50 до 75 м2). Принятый нами дизайн эксперимента (одна площадка, характеризующая одну микрогруппировку) позволил получить достаточную по размеру выборку для проверки статистических гипотез.
Геоботанические описания на площадках 20×20 м были выполнены по общепринятым методикам (Mucina et al., 2000; Методические подходы…, 2010). Были составлены списки видов растений, включая сосудистые растения и напочвенные мхи, с указанием проективного покрытия каждого вида (в %). Дана оценка проективного покрытия ярусов (древесного, кустарникового, травяного, мохового). Географические координаты (WGS84) местоположения площадок были установлены с использованием навигатора Garmin 64st. Описания на площадках 20×20 м выполнены в летнее время в 2016–2020 гг.
Описания микрогруппировок были выполнены на площадках 5×5 м летом 2019 г. Составлены списки видов (учитывались только виды сосудистых растений) с указанием проективного покрытия каждого вида (в %), проведена оценка проективного покрытия ярусов (древесного, кустарникового, травяного).
Для центральных деревьев были определены следующие параметры: высота дерева и расстояние до нижней живой ветви (при помощи лазерного высотомера Nikon Forestry Pro), диаметр ствола на уровне груди, площадь кроны (по формуле для эллипса через диаметры), протяженность кроны (как разность высоты и расстояния до нижней живой ветки).
Для площадок в программе SpeDiv (Смирнов, 2006) были получены оценки экологических факторов по шкалам Элленберга (освещенность, влажность почвы, реакция почвы, богатство почвы азотом) и доли ЭЦГ (бореальная — Br, луговая — Md, неморальная — Nm, нитрофильная — Nt) (Смирнов и др., 2006).
Дифференциация растительности на уровнях фитоценозов (использованы полные списки видов описаний на площадках 20×20 м) и микрогруппировок (списки видов сосудистых растений травяного яруса на площадках 5×5 м) продемонстрирована методом неметрического многомерного шкалирования (NMDS) с использованием метрики Брея-Кёртиса в пакете vegan в среде R (Oksanen et al., 2019). Интерпретация осей ординации проведена путем установления связей осей с экологическими факторами и долями видов ЭЦГ.
Для определения зависимости видового состава микрогруппировок от следующих факторов: а) типа фитоценоза, б) вида эдификатора, в) площадки 20×20 м был осуществлен ряд проверок статистических гипотез. В качестве нулевой гипотезы H0 всегда предполагалось отсутствие зависимости между фактором (или комбинацией факторов) и видовым составом микрогруппировки. Проверка гипотез производилась при помощи статистических пертурбационных тестов для статистик, представленных соотношением средних расстояний Брея-Кёртиса между видовым составом микрогруппировок внутри групп, заданных значением фактора, и всего массива данных. Реализованное в наблюдаемых данных значение статистики сравнивалось с распределением статистики в рамках нулевой гипотезы, полученным при помощи рандомизации групповой принадлежности микрогруппировки. На основании экстремальности реализованного значения принималось решение об отклонении гипотезы H0 на традиционно рассматриваемых уровнях статистической значимости (p = 0.001, 0.01, 0.05). Данный анализ был выполнен с использованием набора базовых команд и инструментов программной среды R.
Для более детальной количественной оценки взаимосвязи между видовым составом микрогруппировок и различных факторов мы провели статистический анализ с использованием метода моделирования совместного распределения ансамбля видов (JSDM) (Warton et al., 2015). Данный подход совмещает методологию обобщенной линейной регрессии и факторного анализа в рамках единой статистической модели. Из доступного набора численных реализаций JSDM мы использовали пакет Hmsc-R в программной среде R, который позволяет наиболее гибко учитывать специфику структуры наблюдаемых данных, а также использует аппарат байесовской статистики для численного расчета параметров модели (Ovaskainen et al., 2017; Norberg et al., 2019; Tikhonov et al., 2020; Ovaskainen, Abrego, 2020). Построенная статистическая модель позволяет оценить вероятность присутствия/отсутствия видов в микрогруппировках в зависимости от экологических параметров, а также определить структуру межвидовых ассоциаций для разных вариантов объединения микрогруппировок. В качестве экологических параметров, используемых в компоненте линейной регрессии JSDM, были выбраны оценки по шкалам Элленберга по освещенности, увлажнению, кислотности и плодородию почвы.
Индикаторные виды микрогруппировок и комбинаций микрогруппировок были определены с использованием метода IndVal (вариант Multilevel pattern analysis) реализованного в пакете indicspecies в программной среде R (De Cáceres, Legendre, 2009). Индекс IndVal был предложен Dufrêne и Legendre (1997). Он представляет собой комбинацию средних обилий видов и частоты их встречаемости в группах. Высокое индикаторное значение достигается при одновременном выполнении двух условий: высокое среднее обилие в пределах группы по сравнению с другими группами («специфичность», specificity) и присутствие в большинстве описаний этой группы («верность», fidelity). Группы описаний могут быть определены различными способами. В нашей работе они соответствуют типам микрогруппировок, заданных эдификатором. Апостериорная статистическая значимость индикаторных значений видов оценивается с помощью перестановочного теста (Borcard et al., 2011).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Данные по морфометрическим параметрам центральных деревьев микрогруппировок представлены в табл. 1. Самый высокий возраст имели дубы и ели — до 100–115 лет. Возраст лип варьировал от 75 до 95 лет, берез — 75–80 лет. Ели имели самые большие высоты и диаметры стволов. Также у елей отмечена наибольшая протяженность кроны, при этом она довольно высоко поднята: в среднем на 8–10 м от земли. Минимальные значения площади кроны были получены для дубов. Таким образом, можно заключить, что среди исследованных деревьев ель отличается максимальными размерами и возрастом (эти параметры прямо связаны с силой фитогенного поля).
Таблица 1. Параметры эдификаторов микрогруппировок
| Тип фитоценоза | Эдификатор | Число деревьев | Возраст, лет | Диаметр ствола, см | Высота дерева, м | Площадь кроны, м2 | Длина кроны, м |
| Березовый | Б | 3 | 80 | 37.0 ± 9.5 | 21.7 ± 2.3 | 73.1 ± 13.6 | 13.7 ± 3.2 |
| Дубово-еловый | Е | 10 | 100–115 | 57.5 ± 11.6 | 28.7 ± 4.5 | 63.1 ± 23.4 | 19.9 ± 4.4 |
| Д | 10 | 100–115 | 45.2 ± 8.3 | 22.6 ± 1.2 | 55.1 ± 17.4 | 11.9 ± 3.5 | |
| Лп | 3 | 80 | 44.7 ± 7.5 | 24.0 ± 1.0 | 58.6 ± 20.3 | 15.0 ± 1.0 | |
| Липовый | Лп | 6 | 90–95 | 49.8 ± 7.9 | 22.5 ± 0.8 | 71.7 ± 27.7 | 8.8 ± 3.1 |
| Березово-липовый | Б | 6 | 75 | 43.5 ± 8.6 | 24.5 ± 2.1 | 66.1 ± 21.5 | 12.0 ± 2.3 |
| Лп | 3 | 75 | 39.7 ± 4.5 | 23.7 ± 2.1 | 55.5 ± 12.4 | 9.0 ± 4.4 |
На рис. 1 показана ординация всех описаний площадок 20×20 м, выполненных в пределах Валуевского лесопарка, и отмечены площадки, на которых и в непосредственной близости с которыми были описаны микрогруппировки. Можно увидеть, что в ординационном пространстве площадки образуют два скопления: справа на диаграмме расположены площадки липняков и березово-липовых фитоценозов, слева — площадки дубово-еловых лесов. Флористические различия между этими типами фитоценозов в основном определяются соотношением неморальных и бореальных видов. Площадка Val19-04 — березовый фитоценоз, по флористическому составу ближе к площадкам дубово-еловых фитоценозов, чем к площадкам липовых и березово-липовых фитоценозов. Но по сравнению с другими типами для березового фитоценоза характерно увеличение доли нитрофильных и луговых видов, а также более высокие значения факторов освещенности и влажности почвы. Ранее было показано, что в зависимости от предшествующей истории природопользования в пределах Москворецко-Окской равнины формируются различные типы березняков (Коротков, 1999). Так как было установлено, что по флористическому составу описания липовых и березово-липовых фитоценозов очень близки между собой (что было ожидаемо, т. к. эти типы представляют последовательные сукцессионные стадии (Аккумуляция…, 2018)), в анализе микрогруппировок они рассматривались как объединенный тип фитоценозов.
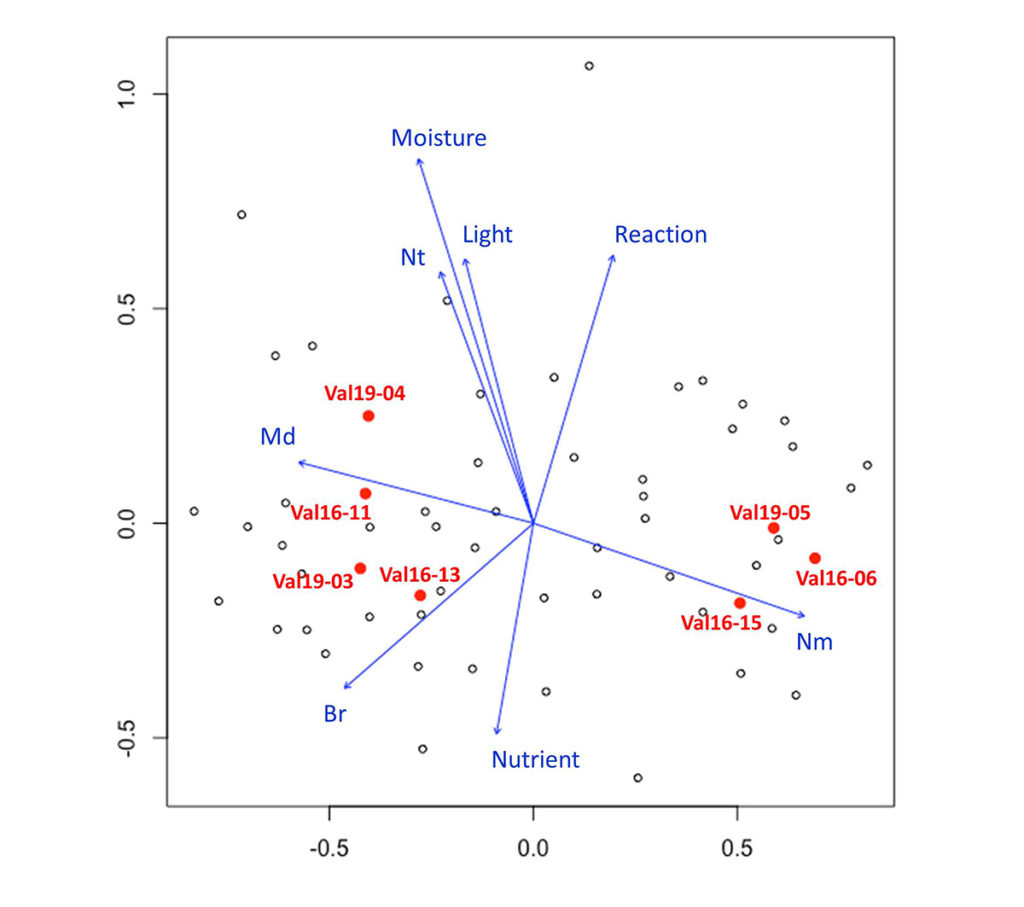
Рисунок 1. Ординация методом NMDS площадок 20×20 м
Примечания: Красными точками обозначены и подписаны 7 площадок, на которых были выполнены описания микрогруппировок.
Микрогруппировки на диаграмме NMDS-ординации (рис. 2) образуют скопления, отражающие дифференциацию растительности на уровне типа фитоценоза (дубово-еловый, березово-липовый, березовый). Первая ось ординации связана с соотношением неморальных и бореальных видов: микрогруппировки с абсолютным доминированием видов неморальной ЭЦГ расположены правее по 1-й оси ординации, а микрогруппировки с большим участием видов бореальной ЭЦГ — левее. Вторая ось ординации связана с долей видов лугово-опушечной и нитрофильной ЭЦГ, по этой оси отчетливо дифференцированы березовые микрогруппировки из березового типа фитоценоза. Отметим, что липовые микрогруппировки из дубово-елового типа имеют большее сходство с еловыми и дубовыми микрогруппировками того же типа фитоценоза, чем с липовыми микрогруппировками из березово-липового типа. Результаты ординации не позволяют уверенно говорить о различиях между микрогруппировками с разными эдификаторами в пределах одного типа фитоценоза.
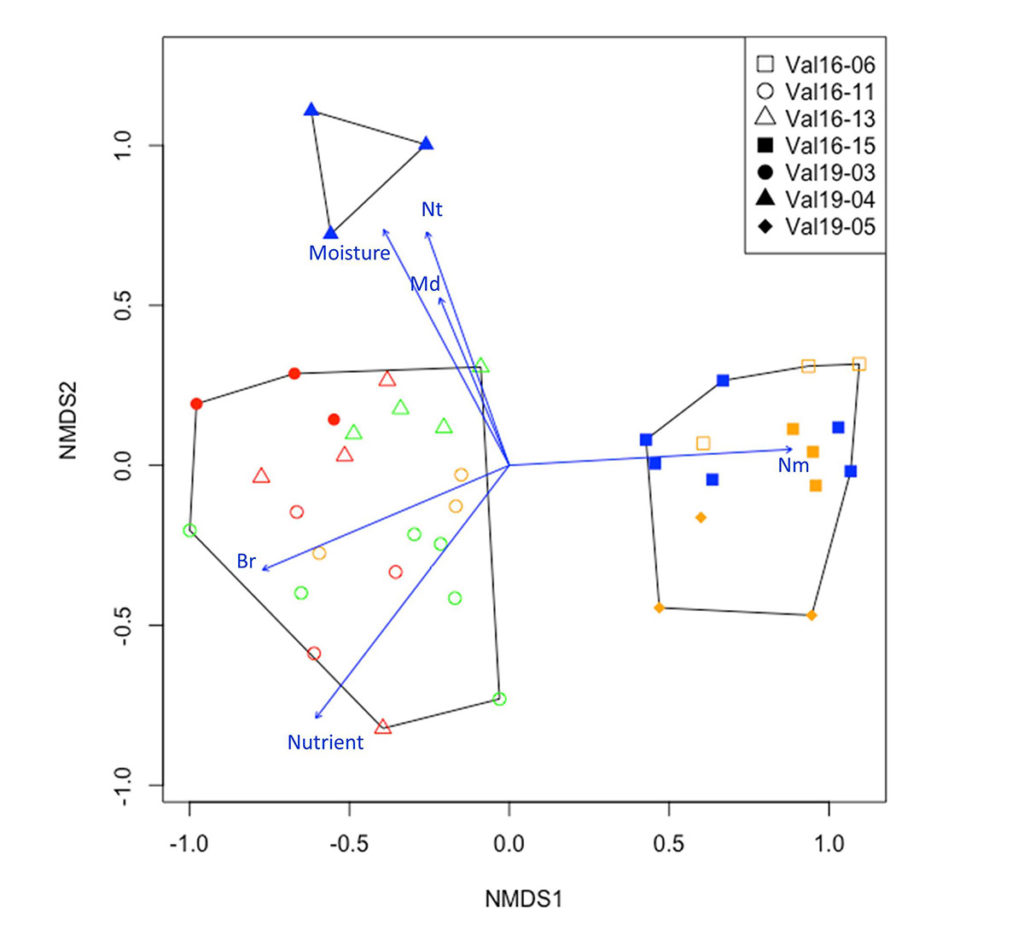
Рисунок 2. Ординация NMDS площадок 5×5 м
Примечания: Цвет значков соответствует породам деревьев: красный — ель, зеленый — дуб, синий — береза, желтый — липа; тип значков обозначает пробные площади 20×20 м, к которым относятся микрогруппировки
Таблица 2. Результаты проверки статистических гипотез
| № | Рассматриваемый экологический вопрос | Формулировка гипотезы H0 | Результат статистического теста |
| 1 | Микрогруппировки с одинаковым эдификатором имеют большее сходство между собой, чем с микрогруппировками с разными эдификаторами | Попарное расстояние между микрогруппировками не зависит от вида эдификатора | H0 отклонена с p-value 0.001 |
| 2 | Микрогруппировки из одного типа фитоценоза имеют большее сходство между собой, чем с микрогруппировками из других типов фитоценозов | Попарное расстояние между микрогруппировками не зависит от типа фитоценоза | H0 отклонена с p-value 0.001 |
| 3 | Микрогруппировки, расположенные близко друг от друга (на одной площадке 20×20 м) имеют большее сходство, чем микрогруппировки, расположенные на разных площадках 20×20 м | Попарное расстояние между микрогруппировками не зависит от того на каких площадках 20×20 м они расположены | H0 отклонена с p-value 0.001 |
| 4 | В пределах одного типа фитоценоза сходство микрогруппировок с одинаковым эдификатором выше, чем сходство микрогруппировок с разными эдификаторами | Попарное расстояние между микрогруппировками в одном типе фитоценоза не зависит от вида эдификатора | H0 отклонена с p-value 0.05, но не может быть отклонена с p-value 0.01 |
| 5 | В пределах одной площадки 20×20 м сходство микрогруппировок с одинаковым эдификатором выше, чем сходство микрогруппировок с разными эдификаторами | Попарное расстояние между микрогруппировками в пределах одной площадки 20×20 м не зависит от вида эдификатора | H0 не может быть отклонена с p-value 0.05 |
| 6 | Сходство микрогруппировок с одним эдификатором в одном типе фитоценоза выше, чем сходство микрогруппировок с одним эдификатором в разных типах фитоценозов | Попарное расстояние между микрогруппировками с одним видом эдификатора не зависит от типа фитоценоза | H0 отклонена с p-value 0.001 |
| 7 | Сходство микрогруппировок в дубово-еловом фитоценозе ниже, чем сходство микрогруппировок в липово-березовом фитоценозе | Попарное расстояние между микрогруппировками не зависит от того, находятся ли они в дубово-еловом или в липово-березовом фитоценозе | H0 отклонена с p-value 0.001 |
Проведенные проверки статистических гипотез о связи видового состава микрогруппировок (табл. 2) позволяют сделать следующие выводы:
- Установлена значимая связь видового состава микрогруппировок с типом фитоценоза.
- Установлена значимая связь видового состава микрогруппировок с площадкой 20×20 м, что можно объяснить зависимостью расселения видов от расстояния, а также тем, что в пределах небольших участков сохраняются выровненные экотопические условия и одинаковая история природопользования (в первую очередь это различные виды лесохозяйственных мероприятий).
- Достоверность различий микрогруппировок разных эдификаторов внутри типа фитоценоза получила ограниченное подтверждение. При этом достоверность различий микрогруппировок разных эдификаторов внутри площадки 20×20 м не получила подтверждения.
- Микрогруппировки с одним эдификатором в разных типах фитоценозов имеют меньшее сходство между собой, чем с микрогруппировками из «своего» фитоценоза. Выводы 3 и 4 говорят о слабом влиянии одиночных деревьев-эдификаторов на видовой состав микрогруппировок.
- Видовой состав микрогруппировок в типе березово-липового фитоценоза более выровненный по сравнению с видовым составом микрогруппировок в типе дубово-елового фитоценоза. Этот факт может быть связан с несколькими причинами, в т. ч. с большим разнообразием (видовое богатство, более сложная эколого-ценотическая структура) дубово-елового типа фитоценоза по сравнению с березово-липовым (меньшее видовое богатство, абсолютное доминирование видов неморальной ЭЦГ) (Носова и др., 2005), а также различиями в парцеллярной структуре этих типов фитоценозов (Комаров и др., 2021).
Как показали результаты моделирования (рис. 3), фактор освещенности не влияет на присутствие/отсутствие видов в микрогруппировках. Это можно объяснить тем, что в подкроновом пространстве условия освещенности не сильно различаются, а также недостаточной чувствительностью шкал Элленберга для выявления небольших различий. Установлена отрицательная связь видов из бореальной и нитрофильной ЭЦГ с фактором реакции почвы и положительная — со многими неморальными видами (исключением являются Dryopteris carthusiana и Paris quadrifolia).
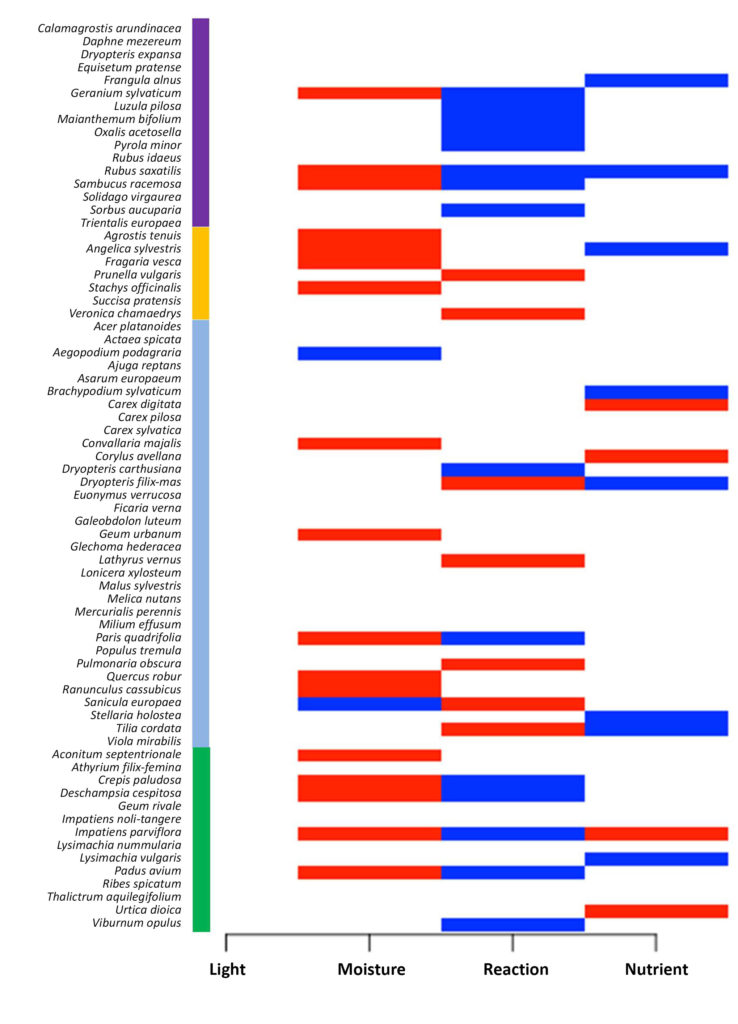
Рисунок 3. Влияние экологических факторов на видовое распределение в JSDM
Примечания: Цветовая шкала слева показывает принадлежность видов к ЭЦГ: фиолетовый — бореальная, желтый — луговая, голубой — неморальная, зеленый — нитрофильная. Пары вид–фактор, для которых была определена статистически значимая связь на уровне, соответствующем апостериорной вероятности 95%, отображены красным (положительные связи) или синим (отрицательные связи), остальные пары отображены белым цветом.
Значимая ассоциированность видов после учета влияния экологических факторов была оценена на уровнях типа фитоценоза и площадок 20×20 м. Результаты анализа представлены на рис. 4 и 5. На уровне типа фитоценоза положительно связаны друг с другом (т. е. частота совместной встречи этих видов в определенном типе фитоценоза выше случайной) 8 видов — все они характерны для дубово-елового типа фитоценоза. Отрицательно связан с этой группой только один вид — Mercurialis perennis L., он распространен в березово-липовых фитоценозах. На уровне площадки (после учета ассоциированности на уровне фитоценозов) выявлена положительная связь между группой, состоящей в основном из видов неморальной группы, с одной стороны, и группой видов, представляющих бореальную и лугово-опушечную ЭЦГ, в нее же попала Lonicera xylosteum L. Эти ассоциативные связи проявляются на локальном уровне и, вероятно, связаны с сукцессионными процессами.
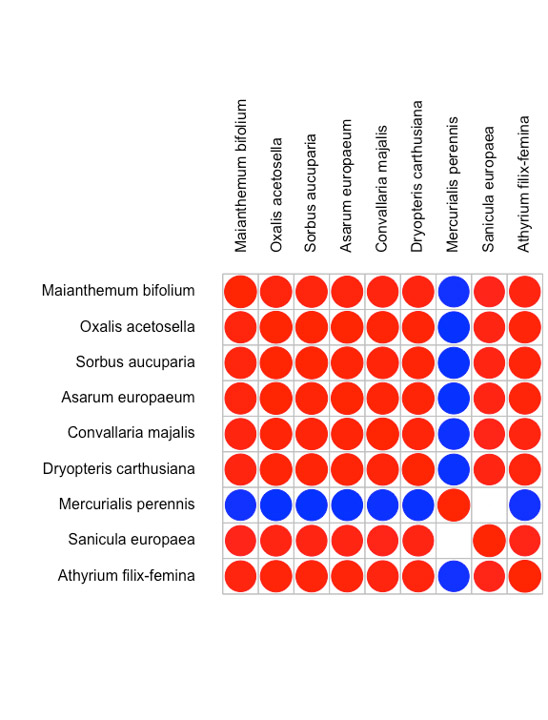
Рисунок 4. Ассоциированность видов на уровне типов фитоценозов в JSDM
Примечания: Отображены только те виды сообществ, для которых была определена хотя бы одна статистически значимая ассоциативная связь (на уровне статистической значимости соответствующей апостериорной вероятности 95%).
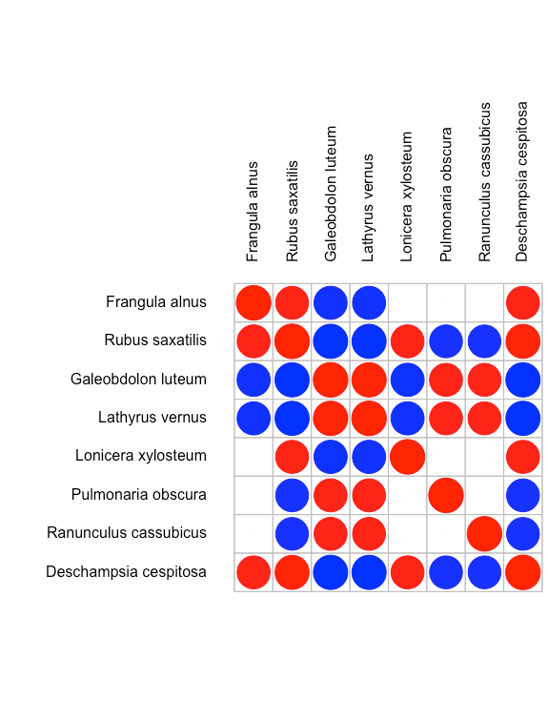
Рисунок 5. Ассоциированность видов на уровне площадок в JSDM
В результате анализа индикаторных видов (табл. 3) было установлено, что липовые и дубовые микрогруппировки не имеют собственных индикаторных видов. Большинство индикаторных видов еловых микрогруппировок относится к бореальной ЭЦГ. Комбинация березовых и липовых микрогруппировок представляет собой тип березово-липового фитоценоза, ее индикаторными видами являются Carex pilosa Scop., Aegopodium podagraria L. и Mercurialis perennis, представляющие неморальную ЭЦГ. Список индикаторных видов для комбинации еловых и дубовых микрогруппировок значительно более длинный, он включает виды разных ЭЦГ, наиболее высокие индикаторные значения среди них имеют Oxalis acetosella L. и Asarum europaeum L. Результаты анализа отражают более сильное эдификаторное влияние ели по сравнению с лиственными деревьями на растительность нижних ярусов, а также более выраженные различия видового состава на уровне типов фитоценозов, чем на уровне микрогруппировок.
Таблица 3. Индикаторные виды микрогруппировок с разными эдификаторами
| Микрогруппировки / комбинации микрогруппировок | Вид | IndVal | p-value |
| Б | Acer platanoides | 0.775 | 0.002 |
| Angelica sylvestris | 0.471 | 0.039 | |
| Е | Rubus saxatilis | 0.670 | 0.010 |
| Maianthemum bifolium | 0.665 | 0.010 | |
| Urtica dioica | 0.586 | 0.010 | |
| Actaea spicata | 0.548 | 0.030 | |
| Luzula pilosa | 0.548 | 0.038 | |
| Б + Лп | Carex pilosa | 0.837 | 0.001 |
| Aegopodium podagraria | 0.777 | 0.002 | |
| Mercurialis perennis | 0.655 | 0.008 | |
| Е + Д | Oxalis acetosella | 0.959 | 0.001 |
| Asarum europaeum | 0.928 | 0.001 | |
| Convallaria majalis | 0.873 | 0.001 | |
| Sorbus aucuparia | 0.828 | 0.009 | |
| Lonicera xylosteum | 0.699 | 0.008 | |
| Corylus avellana | 0.665 | 0.016 | |
| Impatiens parviflora | 0.632 | 0.013 | |
| Athyrium filix-femina | 0.605 | 0.015 |
В нашем исследовании мозаичности лесного покрова мы ставили задачу выявления и оценки различий флористического состава микрогруппировок в разных типах лесных фитоценозов. Обзор литературы позволяет говорить, что наиболее сильное воздействие на подпологовую растительность оказывают деревья-эдификаторы, но их влияние зависит как от климатических и экотопических условий, так и от типов фитоценозов, а также от возрастного и жизненного состояния эдификатора.
Выраженность различий между микрогруппировками может быть связана с климатическими условиями. В исследовании Ястребова и Лычной (1993) показано, что изменения встречаемости вереска и брусники под воздействием фитогенного поля сосны в Карелии выражены гораздо сильнее, чем в Ленинградской области. Авторы делают вывод о том, что «в более суровых экологических условиях даже небольшое воздействие древостоя способно вызвать достаточно сильную реакцию подчиненных ярусов».
Специфичность воздействия деревьев-эдификаторов через формирование фитогенного поля проявляется в «подборе» ими определенных видов-спутников (Маслов, 1990). Известно, что ель оказывает сильное влияние на растительность нижних ярусов посредством изменения параметров верхнего горизонта почвы, связанных с хвойной подстилкой: повышение кислотности, уменьшение содержания оснований, более широкое отношение C/N, накопление подстилки и замедление высвобождения питательных веществ. Это определяет увеличение доли ацидофильных видов, особенно корнеподстилочных, и снижение доли нитрофильных видов (Máliš et al., 2012). Толстый слой подстилки может препятствовать распространению некоторых травянистых видов, в то время как другие виды способны расти в этих условиях (van Oijen et al., 2005).
В нашем случае влияние ели проявляется сильно, но оно заметно на уровне фитоценоза. Небольшие различия между микрогруппировками в пределах дубово-елового типа фитоценоза определяются не отсутствием влияния ели, а слабым эдификаторным влиянием лиственных деревьев (дуба и липы), которое не приводит к сильному изменению растительности нижних ярусов.
Одной из причин слабо выраженных различий между растительным покровом микрогруппировок в многовидовых смешанных лесах может быть влияние соседних микрогруппировок. По оценкам Л. О. Карпачевского (1977), за пределы парцеллы выносится около 30% хвои и листьев. В случае когда граничат парцеллы, сформированные разными эдификаторами, образуется опад смешанного состава, что снижает пространственную контрастность различий параметров верхних горизонтов почвы (Припутина и др., 2020). Кроме деревьев верхнего полога, в формировании смешанного опада участвуют деревья второго подъяруса и кустарники. Было показано (Маслов, 1990), что по влиянию на растительность нижних ярусов рябина, жимолость и лещина очень близки к широколиственным видам деревьев.
А. А. Маслов (1990) установил, что вклад деревьев в формирование мозаичности зависит не только от их эдификаторной силы, но и от размещения особей по площади. Так, мозаичность липово-елового фитоценоза (состав древостоя 8Е2Лп ед. Д, Ос, Б) оказалась связанной не с отдельными деревьями, а с характером размещения достаточно крупных по размеру куртин широколиственных деревьев.
В нашей работе установлено, что различия в растительности намного сильнее выражены на уровне фитоценозов, чем на уровне микрогруппировок. Этот вывод согласуется с результатами многих авторов, полученными ранее. Например, П. Д. Ярошенко (1953, с. 46) описывает исследование микрогруппировок в буковых лесах в окрестностях Львова (Украина), в которых определялись видовой состав, условия освещения, характер подстилки, кислотность почвы и содержание гумуса в почве. Было показано, что «отдельные микрогруппировки одного и того же лесного сообщества хотя и различаются между собой по всем этим признакам, но различия эти менее устойчивы, чем различия между разными лесными ассоциациями, т. е. типами самих сообществ».
С. В. Копцик с соавторами (2008) на основе результатов исследования мозаичности почвенного покрова в НП «Смоленское Поозерье» делают вывод, что «продолжительность воздействия отдельных деревьев в течение их жизни недостаточна для трансформации большинства почвенных свойств, отличающихся сравнительной консервативностью». Время существования микрогруппировки, сформированной деревом-эдификатором, ограничено продолжительностью жизни этого дерева (в среднем редко больше 100–150 лет в условиях Центра Русской равнины). В многовидовых лесных сообществах на месте, освободившемся после гибели дерева, может сформироваться микрогруппировка с другим эдификатором.
Л. О. Карпачевский (1977) выявил большие различия в характере гумусированности почв оконных парцелл широколиственно-еловых лесов (исследования проводились на стационаре «Малинки» в пределах Москворецко-Окской равнины). Он отмечает, что кроме состава растительности данной и соседних парцелл, «большое значение имеет возраст окна и особенности предыдущей парцеллы и характер ее смены: осиновая или еловая парцелла была сформирована на месте окна раньше и произошла смена в результате выворота или через стадию пня» (Карпачевский, 1977, с. 88).
Таким образом, можно говорить о том, что современный растительный покров наследует или запечатлевает в структуре и видовом составе предшествующие состояния за длительный период времени. Это хорошо документировано в работах многих авторов, отмечавших сохранение на протяжении столетий различий между лесными сообществами, сформировавшимися на участках с разной историей природопользования (Коротков, 1999; Тихонова и др., 2012).
ВЫВОДЫ
Изучение внутриценотической неоднородности растительного покрова в отечественной геоботанике имеет почти 100-летнюю историю. Но до сих пор остается много вопросов, связанных с понятийным аппаратом данной области знаний. Проанализировав по многочисленным публикациям формулировки и описания основных терминов, предложенные разными авторами, мы пришли к выводу, что в качестве единицы для исследования внутрибиогеоценотической неоднородности можно оставить парцеллу (в понимании Н. В. Дылиса и Л. О. Карпачевского), а при рассмотрении растительного компонента того же пространственного уровня использовать понятие микрогруппировки.
Результаты всех проведенных анализов мозаичности растительного покрова на уровне микрогруппировок в нескольких широко распространенных на Москворецко-Окской равнине типах лесных фитоценозов показывают, что дифференциация растительности намного сильнее выражена на уровне фитоценозов, чем на уровне микрогруппировок. Среди рассмотренных деревьев-эдификаторов (ель, дуб, береза, липа) наиболее сильное влияние на растительность нижних ярусов оказывает ель, этот вывод согласуется с имеющимися в литературе результатами.
Слабая дифференциация растительности на уровне микрогруппировок в рассмотренных типах елово-широколиственных фитоценозов может определяться рядом причин: влиянием соседних микрогруппировок; недостаточной для заметной трансформации растительности нижних ярусов продолжительности воздействия отдельных деревьев; наследованием в структуре и видовом составе предшествующих состояний за длительный период времени.
БЛАГОДАРНОСТИ
Авторы благодарят М. В. Семенцову, А. Ю. Захаринского, И. М. Аверченкова, Е. В. Ручинскую, Е. В. Басову, А. В. Комарова, а также студентов МГУ и РУДН за участие в полевых исследованиях. Выражаем искреннюю признательность Ю. А. Насимовичу за консультации по флоре Валуевского лесопарка. Исследование выполнено в рамках темы государственного задания ЦЭПЛ РАН № АААА-А18-118052590019-7, полевые работы проведены за счет средств проекта Российского научного фонда № 16-17-10284).
СПИСОК ЛИТЕРАТУРЫ
Аккумуляция углерода в лесных почвах и сукцессионный статус лесов / Н. В. Лукина (ред.). М: Т-во науч. изд. КМК, 2018. 232 с.
Ашик Е. В., Тиходеева М. Ю. Исследование фитогенного поля Tilia cordata Mill. в посадках Pinus silvestris L. в заповеднике «Белогорье» Белгородской области // Вестник СПбГУ. Сер. 3. 2006. Вып. 3. С. 64–73.
Дылис Н. В. Структура лесного биогеоценоза. М.: Наука, 1969. 55 с.
Заугольнова Л. Б. Современные представления о структуре растительного покрова: концепция иерархического континуума // Успехи современной биологии. 1999. Т. 119. № 2. С. 115–127.
Ипатов В. С. Фитогенные поля одиночных деревьев некоторых пород в одном экотопе // Бот. журнал. 2007. Т. 92. № 8. С. 1186–1192.
Ипатов В. С., Журавлева Е. Н., Лебедева В. Х., Тиходеева М. Ю. Фитогенное поле Picea abies, P. obovata (Pinaceae) // Ботанический журнал. 2009. Т. 94. № 4. С. 558–568.
Ипатов В. С., Кирикова Л. А. К характеристике фитогенного поля Picea abies (Pinaceae) в зеленомошных сосняках // Бот. журн. 2001. Т. 86. № 5. С. 94–103.
Карпачевский Л. О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во Моск. ун-та, 1977. 312 с.
Карпачевский Л. О., Воронин А. Д., Дмитриев Е. А., Строганова М. Н., Шоба С. А. Почвенно-биогеоценотические исследования в лесных биогеоценозах. М.: Изд-во Моск. ун-та, 1980. 160 с.
Карпачевский Л. О., Зубкова Т. А., Ташнинова Л. Н., Руденко Р. Н. Почвенный покров и парцеллярная структура лесного биогеоценоза // Лесоведение. 2007. № 7. С. 36–42.
Киричок Е. И., Истомина И. И., Копцева Н. С. Фитогенное поле ели в сосняках-зеленомошниках Неруссо-Деснянского полесья // Фундаментальные и прикладные проблемы ботаники в начале XXI века. 2008. Ч. 5. С. 140–142.
Комаров А. В., Ершов Д. В., Тихонова Е. В. Информативность спектральных и морфометрических признаков оконной структуры полога древостоя на основе спутниковых данных // Лесоведение. 2021. № 3. С. 227–239.
Копцик С. В., Копцик Г. Н., Ермаков И. В., Ливанцова С. Ю. Пространственная изменчивость свойств почв в биогеоценозах еловых лесов // Летопись природы 2007. Кн. 1. п. Пржевальское, 2008. С. 36–41.
Коротков В. Н. Природно-исторический заповедник-леспаркхоз «Горки» / О. В. Смирнова, Е. С. Шапошников (ред.). Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия. Санкт-Петербург: РБО, 1999. С. 106–150.
Корчагин А. А. Строение растительных сообществ // Полевая геоботаника. Методическое руководство. Т. 5. Л.: Издательство Академии Наук СССР, 1976. 313 с.
Котов С. Ф. Количественная оценка эдификаторной роли древесных видов // Бот. журнал. 1983. Т. 68. № 1. С. 39–48.
Крышень А. М. К методике изучения фитогенных полей деревьев // Бот. журнал. 1998. Т. 83. № 10. С. 133–142.
Курнаев С. Ф. Дробное лесорастительное районирование Нечерноземного центра. М.: Наука, 1982. 120 с.
Ландшафты Московской области и их современное состояние / И. И. Мамай (ред.). Смоленск: Изд-во СГУ, 1997. 296 с.
Лебедева В. Х., Тиходеева М. Ю., Ипатов В. С. Сравнительная оценка влияния деревьев на напочвенный покров в березняках черничных // Ботанический журнал. 2007. Т. 92. № 5. С. 681–701.
Леса Южного Подмосковья. М.: Наука, 1985. 281 с.
Маслов А. А. Количественный анализ горизонтальной структуры лесных сообществ. М.: Наука, 1990. 160 с.
Маслов А. А. О взаимодействии фитогенных полей деревьев в сосняке чернично-брусничном // Бот. журнал. 1986. Т. 71. № 12. С. 1646–1652.
Материалы лесоустройства, Федеральная служба лесного хозяйства Российской Федерации. Лесоустроительное предприятие. Экспедиция. Таксационное описание (по состоянию на 01.01.05 года). Лесхоз Экспериментальный Москворецкий. 2005.
Методические подходы к экологической оценке лесного покрова в бассейне малой реки / Л. Б. Заугольнова, Т. Ю. Браславская (отв. ред.). М.: Товарищ-во. научн. изданий КМК, 2010. 240 с.
Мучник Е. Э., Тихонова Е. В., Аверченков И. М., Неслуховский И. Ю., Захаринский А. Ю., Комаров А. В., Кожин М. Н., Семенцова М. В. Валуевский лесопарк как перспективная особо охраняемая природная территория в пределах Новой Москвы // Труды Карельского научного центра Российской академии наук. 2020. № 8. С. 90–103.
Носова Л. М., Тихонова Е. В., Леонова Н. Б. Воздействие деревьев-эдификаторов на биологическое разнообразие лесных экосистем // Лесоведение. 2005. № 4. С. 40–48.
Петров В. В., Кузенкова Л. Я. О западной границе геоботанического района широколиственных лесов в Московской области // Вестн. МГУ, сер. биол., почвовед. 1968. № 3. С. 59–69.
Платонова Е. А., Лантратова А. С., Голубин К. Ю. Фитоценотическая роль липы сердцелистной (Tilia cordata Mill.) на природных и урбанизированных территориях // Hortus botanicus. 2006. URL: http://hortus.karelia.ru/bgm/hb.htm (дата обращения 25.07.2021).
Припутина И. В., Фролова Г. Г., Шанин В. Н., Мякшина Т. Н., Грабарник П. Я. Распределение органического вещества и азота в дерново-подбурах Приокско-Террасного заповедника и его связь со структурой лесных фитоценозов // Почвоведение. 2020. № 8. С. 921–933.
Публичная кадастровая карта Московской области 2020. Интернет ресурс. URL: https://публичная-кадастровая-карта.рф/ (дата обращения 09.01.2021).
Самойлов Ю. И., Тархова Т. Н. Анализ фитогенных полей одиночных деревьев с использованием метода главных компонент // Ботанический журнал. 1993. Т. 78. № 5. С. 61–77.
Смирнов В. Э. SPEDIV — программа для анализа разнообразия растительности // В сборнике: Принципы и способы сохранения биоразнообразия / Л. А. Жукова (отв. ред.). Йошкар-Ола: Мар. гос. ун-т, 2006. С. 142–143.
Смирнов В. Э., Ханина Л. Г., Бобровский М. В. Обоснование системы эколого-ценотических групп видов растений лесной зоны Европейской России на основе экологических шкал, геоботанических описаний и статистического анализа // Бюлл. МОИП. Сер. Биологическая. 2006. Т. 111. № 2. С. 36–47.
Сочава В. Б. Классификация растительности как иерархия динамических систем // Геоботаническое картографирование. 1972. С. 3–18.
Справочная информация о погоде и климате. URL: http://meteo.ru (дата обращения: 12.01.2021).
Тарасова Ю. В., Матвеев И. В., Орешкин Д. Г. Влияние одиночных берез (Betula pendula Roth) на характеристики почв некоторых растительных сообществ таежной зоны // Вестник СПбГУ. Сер. 3. 2003. Вып. 3 (№ 19). С. 37–46.
Тихонова Е. В., Пестерова О. А., Семенищенков Ю. А. Синтаксономический анализ искусственных лесов юго-западного Подмосковья // Известия Самарского научного центра Российской академии наук. 2012. Т. 14. № 1-4. С. 1135–1138.
Уранов А. А. Фитогенное поле // Проблемы современной ботаники. М.–Л.: Наука, 1965. Т.1. С. 251–254.
Черняева Е. В., Викторов В. П. История и современное состояние изучения фитогенных полей // Социально-экологические технологии. 2016. № 1. С. 89–106.
Юрков Д. Е., Гаврилина И. А., Неслуховский И. Ю. Филимонковское поселение. Новые районы Москвы. 2019. 280 с.
Ярошенко П. Д. Геоботаника. 1969. М.: Просвещение, 1969. 200 с.
Ярошенко П. Д. К изучению горизонтального расчленения растительного покрова // Ботан. журн. 1958. Т. 43. № 3. С. 381–387.
Ярошенко П. Д. Основы учения о растительном покрове. М.: Географгиз, 1953. 354 с.
Ястребов А. Б., Лычная Н. В. Исследование фитогенных полей деревьев в лишайниково-зеленомошных сосняках // Ботан. журн. 1993. 78. № 5. С. 78–92.
Borcard D., Gillet F., Legendre P. Numerical Ecology with R. New York, NY: Springer Verlag, 2011. 306 p.
De Cáceres M., Legendre P. Associations between species and groups of sites: indices and statistical inference // Ecology. 2009. Vol. 90 (12). P. 3566–3574.
Dufrêne M., Legendre P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach // Ecological Monographs. 1997. Vol. 67. P. 345–366.
Máliš F., Ujházy K., Vodálová A., Barka I., Čaboun V., Sitková Z. The impact of Norway spruce planting on herb vegetation in the mountain beech forests on two bedrock types // European Journal of Forest Research 2012. Vol. 131. P. 1551–1569.
Mucina L., Schaminee J. H. J., Rodwell J. S. Common data standards for recoding releves in field survey for vegetation classification // Journal of Vegetation Science. 2000. Vol. 11. P. 769–772.
Norberg A., Abrego N., Blanchet F. G., Adler F., Anderson B.… & Ovaskainen O. A comprehensive evaluation of predictive performance of 33 species distribution models at species and community levels // Ecological Monographs. 2019. Vol. 89. Article e01370.
Økland R. H., Rydgren K., Økland T. Single-tree influence on understorey vegetation in a Norwegian boreal spruce forest // Oikos. 1999. Vol. 87. P. 488–498.
Oksanen J. F., Blanchet G., Friendly M., Kindt R., Legendre P. … & Wagner H. Vegan: Community Ecology Package. R package version 2.5-6. 2019. URL: https:// CRAN.R-project.org/ package=vegan (дата обращения 09.01.2021).
Ovaskainen O., Abrego N. Joint Species Distribution Modelling (With Applications in R). Cambridge University Press. 2020. 388 p.
Ovaskainen O., Tikhonov G., Norberg A., Blanchet F. G., Duan L., Dunson D., Roslin T. Abrego N. How to make more out of community data? A conceptual framework and its implementation as models and software // Ecology Letters. 2017. Vol. 20. P. 561–576.
Tikhonov G., Opedal Ø. H., Abrego N., Lehikoinen A., de Jonge M. M., Oksanen J., Ovaskainen O. Joint species distribution modelling with the R-package Hmsc // Methods in Ecology and Evolution, 2020. Vol. 11. No 3. P. 442–447.
Van Oijen D., Feijen M., Hommel P., den Ouden J., de Waal R. Effects of tree species composition on within-forest distribution of understorey species // Applied Vegetation Science. 2005. Vol. 8 (2). P. 155–166
Warton D. I., Blanchet F. G., O’Hara R. B., Ovaskainen O., Taskinen S., Walker S. C., Hui F. K. C. So Many Variables: Joint Modeling in Community Ecology // Trends in Ecology & Evolution. 2015. Vol. 30. Iss. 12. P. 766–779.
Zinke P. J. The pattern of influence of individual forest trees on soil properties // Ecology. 1962. Vol. 43. P. 130–133.
Рецензенты: д. б. н. Маслов А. А., д. б. н. Семенищенков Ю. А.





