- DOI 10.31509/2658-607x-2020-3-2-1-20
- УДК 592:574.42
Влияние дождевых червей разных морфо-экологических групп на аккумуляцию углерода в лесных почвах
![]() © 2020 г.
© 2020 г.
А.П. Гераськина
ФГБУН Центр по проблемам экологии и продуктивности лесов Российской академии наук Россия, 117997 Москва, ул. Профсоюзная, 84/32, стр. 14
E-mail: angersgma@gmail.com
Поступила в редакцию 25.05.2020 г.
Принята к печати 12.06.2020 г.
К настоящему времени в лесной экологии не сделаны однозначные выводы о влиянии деятельности таких крупных беспозвоночных-сапрофагов, как дождевые черви, на динамику углерода в почвах. Ряд авторов утверждает, что аккумуляция углерода снижается в результате деятельности дождевых червей. Другие исследования показывают, что дождевые черви способствуют аккумуляции углерода в почвах. При этом в большинстве работ не принимаются во внимание отличия трофической и роющей деятельности отдельных морфо-экологических групп дождевых червей в разных почвенных горизонтах. Цель данной работы: провести дифференцированную оценку влияния дождевых червей разных морфо-экологических групп на аккумуляцию углерода и связанных с ней почвенных параметров (содержание азота и соотношения С/N) в ходе изменения сукцессионного статуса лесов. Полевые работы проведены в весенне-летние периоды 2016 и 2018 гг. в трех регионах: Брянская область (Брянское полесье), Московская область (Москворецко-Окская равнина, Валуевский лесопарк) и Северо-Западный Кавказ (Краснодарский край, Апшеронское лесничество; республика Адыгея, Кавказский биосферный заповедник). В каждом регионе выделяли три основных стадии восстановления хвойно-широколиственного леса после сплошных рубок. Для каждой стадии подобраны по три пробные площади 50х50 м, на которых выполнены геоботанические, почвенные описания и учеты дождевых червей. Установлено, что в ходе изменения сукцессионного статуса лесов происходит усложнение видового состава и набора морфо-экологических групп дождевых червей, но не последовательная замена одних групп другими. Выявлены неоднозначные влияния разных морфо-экологических групп дождевых червей на аккумуляцию углерода в лесных почвах. Установлены отрицательные корреляционные зависимости между суммарной биомассой подстилочных, почвенно-подстилочных и норных видов червей и содержанием углерода в подстилке. При этом биомасса почвенно-подстилочных видов положительно связана с содержанием углерода в гумусовом горизонте. Показатели С/N и содержание азота однонаправленно коррелируют с биомассой дождевых червей в горизонтах их активности: при увеличении биомассы червей разных морфо-экологических групп соотношение С/N уменьшается, содержание азота повышается.
Ключевые слова: тип леса, сукцессионный статус, хроносерия, подстилка, азот, соотношение C/N, беспозвоночные-сапрофаги, биомасса
Продуктивность лесных экосистем во многом определяется активностью почвенных сапрофагов, поскольку основная часть потока энергии направлена по детритной пищевой цепи (Стриганова, 2012). Мощность потока мертвого органического вещества, поступающего в почву, достигает не менее 95% от общего количества органического вещества, ассимилированного продуцентами (Begon et all., 1986). В смешанных и лиственных лесах Европейской части России главными агентами разложения растительного опада выступают крупные сапрофаги, до 90% биомассы которых могут составлять дождевые черви (Абатуров, 1976; Стриганова, 1980).
С одной стороны, растительный опад (листовой, стволовой, корневой) и корневые выделения растений служат источником углерода для дождевых червей (Гончаров, 2014; Gleixner 2013). С другой стороны, дождевые черви обеспечивают закрепление углерода в почве двумя основными путями: гумусообразованием в результате трофической деятельности и переносом углерода с поверхности почвы в нижние горизонты в процессе их активной роющей деятельности. При отсутствии дождевых червей и других крупных сапрофагов деструкция растительного опада осуществляется микроорганизмами и сапротрофной мезо- и микрофауной, которая не способна к переносу углерода в минеральные горизонты почв. Кроме того, в процессе микробного дыхания происходит эмиссия углекислого газа, что ведет к потерям углерода, а не закреплению его в почве (Frouz et all., 2013).
В современных исследованиях глобальной динамики углерода в наземных экосистемах приводятся противоречивые оценки влияния дождевых червей на аккумуляцию углерода в почве. Ряд авторов утверждает, что аккумуляция углерода снижается в результате деятельности дождевых червей (Alban, Berry, 1994; Burtelow et al., 1998; Bohlen et al., 2004). Другие исследования показывают, что дождевые черви способствуют аккумуляции углерода в почвах (Pulleman et al., 2005; Novara et al., 2015). Вероятно, такие противоположные заключения связаны с тем, что в большинстве работ комплекс дождевых червей рассматривают как единое целое, и исследователи учитывают суммарную биомассу всех видов дождевых червей. При этом не принимают во внимание различия отдельных морфо-экологических групп дождевых червей, среди которых по отношению к органическому веществу почв выделяют первичных гумусообразователей – виды, питающиеся на поверхности (подстилочные, почвенно-подстилочные и норные) и вторичных гумусообразователей – виды, питающиеся в почве (собственно почвенные) (Перель, 1979).
Цель данной работы заключалась в дифференцированной оценке влияния дождевых червей разных морфо-экологических групп на аккумуляцию углерода в лесных почвах в ходе изменения сукцессионного статуса лесов.
Задачи:
- исследовать видовой состав, биомассу и динамику морфо-экологических групп дождевых червей в ходе преобразования лесных сообществ;
- проaнализировать влияние дождевых червей 4-х морфо-экологических групп на запасы подстилки и содержание углерода в почвенных горизонтах;
- оценить влияние дождевых червей разных морфо-экологических групп на соотношение С/N и содержание азота, как наиболее значимые почвенные показатели, связанные с процессами аккумуляции углерода, которые регулируются дождевыми червями.
МАТЕРИАЛ И МЕТОДЫ
Полевые работы проведены в весенне-летние периоды 2016 и 2018 гг. в хвойно-широколиственных лесах трех регионов Европейской части России: Брянское Полесье (территория заповедника «Брянский лес» – 52.5464 N, 34.0797 E), Москворецко-Окская равнина (Валуевский лесопарк – 55.5780 N, 37.3272 Е), Северо-Западный Кавказ (Апшеронское лесничество пос. Отдаленный Краснодарского Края – 44.0669 N, 39.7164 Е и кордон Гузерипль Кавказского биосферного заповедника, республика Адыгея – 44.0002 N, 40.1421 E). В каждом регионе обследовали по три типа леса, которые представляют собой различные этапы сукцессионной смены растительных сообществ (хроносерии) после сплошных рубок (Лукина и др., 2018).
В Брянском Полесье исследовали: сосняки кустарничково-зеленомошные бореальнотравные (возраст 40-60 лет), смешанные леса с сосной, елью, дубом, кленом неморально-бореальнотравные или сосняки сложные волосисто-осоково-разнотравные (возраст 70-120 лет), широколиственные леса с елью неморальнотравные (возраст более 120 лет). Тип почв: дерново-подзолистые иллювиально-железистые песчаные на флювиогляциальных песках (Классификация почв…, 2004) или Podzols Albic (WRB, 2015). Содержание физической глины в почвообразующих породах составляет от 0.5 до 5%. Актуальная кислотность почвообразующих пород варьирует от рН 5.1 до 5.7 (Лукина и др., 2018).
На территории Москворецко-Окской равнины исследовали: березово-липовые леса с осиной неморальнотравные (возраст 50-70 лет), липовые леса с березой и осиной неморальнотравные (возраст 90-110 лет), широколиственно-еловые леса бореально-неморальнотравные (возраст 115-125 лет). Почва дерново-подзолистая среднесуглинистая на покровных суглинках, подстилаемых мореной (Классификация почв…, 2004) или Retisols Albic (WRB, 2015). Содержание физической глины в почвообразующих породах варьирует от 34.3 до 45.3%. Актуальная кислотность почвообразующих пород составляет от рН 5.1 до 5.6 (Лукина и др., 2018).
На территории Северо-Западного Кавказа исследовали: осиново-грабовые жимолостно-мелкотравные леса (возраст 45-65 лет), буково-пихтово-грабовые мелкотравные леса (возраст 80-110 лет), пихтово-буковые мертвопокровные леса (возраст более 400 лет). Тип почв: бурозем тяжелосуглинистый на глинистых сланцах (Классификация почв…, 2004) или Cambisols Dystric (WRB, 2015). Содержание физической глины в почвообразующих породах варьирует от 36.5 до 72.7%. Актуальная кислотность почвообразующих пород изменяется от рН 5.5 до 5.7 (Лукина и др., 2018).
Для каждого типа леса в трех регионах подбирали по три пробные площади (ПП) 50х50 м (всего 27), на которых выполняли геоботанические, почвенные описания и учеты дождевых червей (Гераськина, 2018; Лукина и др. 2018; Кузнецова и др., 2019; Шевченко и др., 2019).
Дождевых червей учитывали путем раскопки и ручного разбора почвенных проб: на каждой ПП отбирали от 10 (Брянское полесье, Москворецко-Окская равнина) до 16 проб (Северо-Западный Кавказ) площадью 25х25 см2, глубиной 30 см. Всего в лесах Брянского полесья взято 90 почвенных проб, в лесах Москворецко-Окской равнины – 60, в лесах Северо-Западного Кавказа – 144. Дождевых червей фиксировали в 96% спирте. Видовую идентификацию проводили по определителю Т.С. Всеволодовой-Перель (1997). Морфо-экологические группы дождевых червей классифицировали согласно Т.С. Перель (1979). Биомассу определяли путем взвешивания фиксированных червей с наполненным кишечником.
На каждой ПП под пологом леса закладывали опорные почвенные разрезы, из которых отбирали образцы подстилки (подгоризонт L) и почвы (через каждые 10 см) до почвообразующей породы. Во всех образцах измеряли рН водной вытяжки потенциометрически. Содержание углерода и азота оценивали на CHN анализаторе (ЕА 1110 (CHNS-O) в экоаналитической лаборатории ЦКП «Хроматография» ИБ Коми НЦ УрО РАН. Для определения массы подстилки и запасов углерода в ней на каждой ПП отбирали образцы подстилки 0.25х0.25 м в трехкратной повторности. Рассчитывали запасы углерода в подстилке и минеральных горизонтах почвы согласно Методическим указаниям (2017). Расчеты проводили с учетом реальной мощности горизонтов и для фиксированных слоев 0–30, 0–50, 0–100 см (Лукина и др. 2018; Кузнецова и др., 2019). Выделение почвенных горизонтов: подстилка или органогенный горизонт (О), гумусовый или гумусово-аккумулятивный горизонт (А), элювиальный горизонт (E), иллювиальный горизонт (В), материнская порода (С) проведено согласно Национальному атласу почв Российской Федерации (2011).
Статистическая обработка данных по дождевым червям выполнена с использованием пакетов программ MS Excel 2019 и Statistica 6.0. Для выявления статистически значимых различий применяли непараметрический критерий Краскелла-Уоллиса, уровень значимости (p) принят ≤ 0.05. В связи с тем, что значения биомассы дождевых червей разных морфо-экологических групп сильно варьировали, в дальнейший анализ их связей с аккумуляцией углерода в разных почвенных горизонтах (и сопряженными показателями – содержанием N и соотношением C/N) включали только те морфо-экологические группы дождевых червей, вариация показателя биомассы которых в выборках подчинялись нормальному распределению. На основании значений критерия Пирсона (К набл. < К крит.; К крит. = 12.591) закону нормального распределения соответствовали наборы данных биомассы червей подстилочной группы в лесах Брянского Полесья и Москворецко-Окской равнины, почвенно-подстилочной – в лесах Москворецко-Окской равнины, собственно почвенной – в лесах Северо-Западного Кавказа и Москворецко-Окской равнины, а также суммарная биомасса червей морфо-экологических групп, питающихся на поверхности почвы (подстилочные, почвенно-подстилочные и норные) в лесах Северо-Западного Кавказа и Москворецко-Окской равнины. Указанные ряды данных использовали для линейного регрессионного анализа (коэффициент детерминации R2) с целью выявления корреляционных зависимостей между группами дождевых червей и почвенными параметрами. При этом учитывали горизонты активности разных морфо-экологических групп дождевых червей, на которые черви оказывают существенное влияние. В горизонте подстилки активны подстилочные, почвенно-подстилочные и норные виды, в гумусовом горизонте — почвенно-подстилочные и собственно почвенные виды.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1. Видовое разнообразие и изменение состава морфо-экологических групп дождевых червей в ходе послерубочных сукцессий лесов
1.1 Леса Брянского Полесья
Обнаружены 4 вида дождевых червей двух морфо-экологических групп: подстилочные Dendrodrilus rubidus tenuis (Eisen, 1874), Dendrobaena octaedra (Savigny, 1826); почвенно-подстилочные Lumbricus rubellus Hoffmeister, 1843, Eisenia nordenskioldi (Eisen, 1879) (табл. 1). Всего для зоны хвойно-широколиственных лесов указывается не менее десяти массовых видов (Перель, 1979). Бедный видовой состав в исследованных нами лесах связан, в первую очередь, с легким гранулометрическим составом почв данной территории – во всех трех типах леса почвы супесчаные (Лукина и др., 2018; Кузнецова и др., 2019). Известно, что легкие супесчаные почвы, характеризующиеся слабой влагоудерживащей способностью, неблагоприятны для обитания дождевых червей (Жуков, 2004; Curry, 2004 и др.).
Таблица 1. Видовой состав и численность дождевых червей разных морфо-экологических групп, выявленных в хвойно-широколиственных лесах исследованных регионов
| Морфо-экологическая группа | Регион | Тип леса | Вид | Численность, экз./м2±SE |
| Подстилочные
(4 вида) |
Брянское Полесье | B1 | – | – |
| B2 | D. octaedra | 12.0±4.6 | ||
| D.r. tenuis | 2.0±0.3 | |||
| B3 | D. octaedra | 7.3±2.8 | ||
| D.r. tenuis | 0.7±0.3 | |||
| Всего: 2 вида | ||||
| Москворецко-Окская равнина | M1 | – | – | |
| M2 | D. octaedra | 8.8±0.8 | ||
| L. castaneus | 0.8±0.1 | |||
| M3 | D. octaedra | 6.4±1.3 | ||
| D.r. tenuis | 2.0±0.5 | |||
| L. castaneus | 0.9±0.2 | |||
| Всего: 3 вида | ||||
| Северо-Западный Кавказ | C1 | D. octaedra | 1.3±0.2 | |
| D. attemsi | 0.3±0.1 | |||
| C2 | D. octaedra | 7.0±1.6 | ||
| D. attemsi | 0.7±0.3 | |||
| C3 | D. octaedra | 1.7±0.3 | ||
| D. attemsi | 0.3±0.1 | |||
| D.r. tenuis | 0.3±0.1 | |||
| Всего: 3 вида | ||||
| Почвенно-подстилочные
(3 вида) |
Брянское Полесье | B1 | – | – |
| B2 | E. nordenskioldi | 0.3±0.2 | ||
| B3 | L. rubellus | 1.3±0.4 | ||
| Всего: 2 вида | ||||
| Москворецко-Окская равнина | М1 | L. rubellus | 14.0±4.5 | |
| M2 | L. rubellus | 16.0±4.5 | ||
| M3 | L. rubellus | 42.0±7.6 | ||
| Всего: 1 вид | ||||
| Северо-Западный Кавказ | C1 | E. fetida | 0.5±0.06 | |
| C2 | – | – | ||
| C3 | E. fetida | 0.5±0.1 | ||
| Всего: 1 вид | ||||
| Собственно почвенные
(6 видов) |
Брянское Полесье | B1 | – | – |
| B2 | – | – | ||
| B3 | – | – | ||
| Москворецко-Окская равнина | M1 | А. c. caliginosa | 29.0±4.7 | |
| A. rosea | 32.0±5.6 | |||
| O. lacteum | 4.0±0.5 | |||
| M2 | А. c. caliginosa | 39.2±7.1 | ||
| A. rosea | 16.0±3.9 | |||
| M3 | А. c. caliginosa | 8.0±2.4 | ||
| A. rosea | 4.8±0.9 | |||
| Всего: 3 вида | ||||
| Северо-Западный Кавказ | C1 | A. jassyensis | 11.3±3.0 | |
| D. s. schmidti | 17.7±2.9 | |||
| D. tellermanica | 5.7±1.5 | |||
| C2 | A. jassyensis | 6.3±3.5 | ||
| D. s. schmidti | 22.0±4.8 | |||
| C3 | A. jassyensis | 8.0±1.0 | ||
| D. s. schmidti | 24.6±8.6 | |||
| Всего: 3 вида | ||||
| Норные
(3 вида) |
Брянское Полесье | B1 | – | – |
| B2 | – | – | ||
| B3 | – | – | ||
| Москворецко-Окская равнина | M1 | – | – | |
| M2 | A. longa | 3.2±0.2 | ||
| L. terrestris | 1.6±0.1 | |||
| M3 | A. longa | 1.6±0.4 | ||
| L. terrestris | 0.8±0.3 | |||
| Всего: 2 вида | ||||
| Северо-Западный Кавказ | C1 | D. mariupolienis | 0.5±0.3 | |
| C2 | D. mariupolienis | 2.0±0.2 | ||
| C3 | D. mariupolienis | 6.7±3.3 | ||
| Всего: 1 вид |
Примечание:
В1 – сосняки кустарничково-зеленомошные бореальнотравные,
B2 – смешанные леса с сосной, елью, дубом и др. неморально-бореальнотравные,
B3 – широколиственные леса с елью неморальнотравные
M1 – березово-липовые леса с осиной неморальнотравные,
M2 – липовые леса с березой и осиной неморальнотравные,
M3 – широколиственно-еловые леса бореально-неморальнотравные
С1 – осиново-грабовые жимолостно-мелкотравные леса,
С2 – буково-пихтово-грабовые мелкотравные леса,
С3 – пихтово-буковые мертвопокровные леса
В сосняках кустарничково-зеленомошных бореальнотравных (В1) дождевые черви не обнаружены в почвенных пробах. Кроме того, дождевые черви не найдены в этих лесах и в ходе обследования благоприятных мест обитания: понижений, западин, валежа поздних стадий разложения. Отсутствие червей в этих лесах связано не только с легким гранулометрическим составом почв, но и с качеством опада. В подстилке преобладает трудноразлагаемый опад растений древесного полога (сосна обыкновенная) и напочвенного покрова (черника обыкновенная, брусника обыкновенная, вереск обыкновенный). Кислотность подстилки варьирует от 4.3 до 4.7, что также неблагоприятно для дождевых червей. Оптимальная кислотность для активной жизнедеятельности, в том числе репродукции большинства видов дождевых червей составляют от pH 5.5 до значений близких к нейтральным (Перель, 1979; Hirth et al., 2009; Moore et al., 2013).
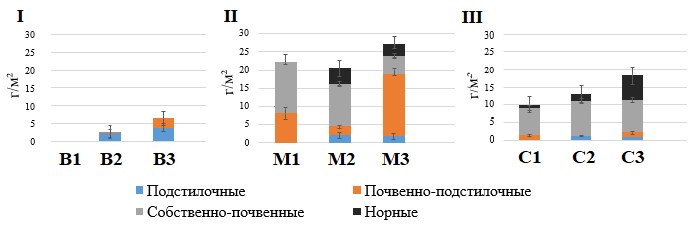
Рисунок 1. Динамика биомассы морфо-экологических групп дождевых червей в ходе изменения сукцессионного статуса лесов
Примечание: I – Брянское Полесье, II – Москворецко-Окская равнина, III – Северо-Западный Кавказ. Обозначение типов леса В1…С3– как и в табл. 1.
В смешанных лесах с сосной, елью, дубом неморально-бореальнотравных (B2, сосняк сложный) обнаружены три вида дождевых червей: подстилочные D. octaedra, D.r. tenuis и почвенно-подстилочный E. nordenskioldi. Численность и биомасса червей низкие (табл. 1, рис. 1), но возможность их обитания в этих лесах в сравнении с сосняками кустарничково-зеленомошными обусловлена наличием легкоразлагаемого опада подроста (липа сердцевидная, клен остролистный) и кустарников (крушина ломкая, лещина обыкновенная). Также подстилка в этих лесах более благоприятна для дождевых червей за счет повышения значений рН до 5.5 до 5.9, в сравнении с предыдущей стадией сукцессии.
В широколиственных лесах с елью неморальнотравных (B3) выявлены три вида люмбрицид: подстилочные D. octaedra, D.r. tenuis и почвенно-подстилочный L. rubellus. Численность и биомасса дождевых червей остаются низкими, как и в сложных сосняках (табл. 1, рис. 1), несмотря на увеличение доли легкоразлагаемого опада древесного яруса (липа сердцевидная, клен остролистный, ясень обыкновенный), подроста (липа, клен, вяз голый) и кустарников (лещина обыкновенная, черемуха обыкновенная). Кислотность подстилки в этих лесах – в пределах оптимальных значений: рН от 5.9 до 6.4, что также не является лимитирующим фактором для обитания дождевых червей.
Выявлены статистически значимые различия в биомассе подстилочных и почвенно-подстилочных видов червей между смешанными лесами с сосной, елью, дубом (В2) и широколиственными лесами с елью (В3), а также различия в общей биомассе червей на трех стадиях сукцессии (рис. 1, табл. 2).
Таблица 2. Значения Н-критерия Краскела-Уоллиса при сравнении биомассы дождевых червей разных морфо-экологических групп в разных типах леса
| Морфо-экологическая группа | Сравниваемые типы леса | df | Нu | Р |
| Подстилочные | *B2 х B3 | 1 | 2.167 | 0.041** |
| M2 х M3 | 1 | 0.079 | 0.777 | |
| C1 х С2 х С3 | 2 | 0.492 | 0.781 | |
| Почвенно-подстилочные | B2 х B3 | 1 | 1.750 | 0.048** |
| M2 х M2 х M3 | 2 | 4.610 | 0.036** | |
| C1 х С3 | 1 | 0.437 | 0.508 | |
| Собственно почвенные | M2 х M2 х M3 | 2 | 8.564 | 0.014** |
| C1 х С2 х С3 | 2 | 0.862 | 0.649 | |
| Норные | M2 х M3 | 1 | 0.784 | 0.375 |
| С2 х С3 | 1 | 4.419 | 0.012** | |
| Все группы | B1 х B2 х B3 | 2 | 6.882 | 0.032** |
| M2 х M2 х M3 | 2 | 0.929 | 0.629 | |
| C1 х С2 х С3 | 2 | 0.965 | 0.617 |
Примечание: *Обозначение типов леса В1…С3 – как и в табл. 1.
**Различия статистически значимы (р≤0.05).
Таким образом, в почвах легкого гранулометрического состава Брянского Полесья, при смене растительных сообществ и изменении качества опада на более благоприятный для дождевых червей, происходит заселение подстилочных и почвенно-подстилочных видов, которые, тем не менее, остаются малочисленными, и их функциональная роль в деструкции подстилки невелика (Лукина и др., 2018).
1.2 Леса Москворецко-Окской равнины
Обнаружены 9 видов дождевых червей, принадлежащих к 4-м морфо-экологическим группам: подстилочные D.r. tenuis, D. octaedra, Lumbricus castaneus (Savigny, 1826); почвенно-подстилочный L. rubellus, собственно почвенные Aporrectodea caliginosa caliginosa (Savigny, 1826), Aporrectodea rosea (Savigny, 1826), Octolasium lacteum (Oerley, 1885); норные Lumbricus terrestris Linnaeus, 1758, Aporrectodea longa (Ude, 1885). Благоприятными факторами для жизнедеятельности дождевых червей в этих лесах служит гранулометрический состав почв (почвы среднесуглинистые), присутствие легкоразлагаемого опада лиственных пород деревьев и кустарников, а также оптимальная кислотность подстилки (5.8-6.1) на всех стадиях сукцессии.
В березово-липовых лесах с осиной неморально-травных (M1) обнаружены 4 вида дождевых червей: почвенно-подстилочный L. rubellus и собственно почвенные A. с. caliginosa, A. rosea и O. lacteum. Наибольший вклад в биомассу вносят собственно почвенные виды (рис. 1). Отсутствие подстилочных видов, вероятно, связано с быстрой утилизацией легкоразлагаемого опада липы и березы почвенной биотой (Березина, 2016).
В липовых лесах с березой и осиной неморальнотравных (M2) выявлены 7 видов дождевых червей: подстилочные D. octaedra, L. castaneus, почвенно-подстилочный L. rubellus, собственно почвенные A. с. caliginosa, A. rosea, норные L. terrestris, A. longa (табл. 1). Несмотря на представленность 4-х морфо-экологических групп люмбрицид, биомасса собственно почвенных видов превышает биомассу червей других групп (рис. 1).
В широколиственно-еловых лесах бореально-неморальнотравных (M3) выявлено 8 видов дождевых червей: подстилочные D. octaedra, D.r. tenuis, L. castaneus, почвенно-подстилочный L. rubellus, собственно почвенные A. caliginosa, A. rosea, норные L. terrestris, A. longa. Отличием широколиственно-елового леса от двух предыдущих стадий хроносерии является существенное снижение биомассы собственно почвенных видов и ее увеличение у почвенно-подстилочного вида L. rubellus в 4.5 раза.
Таким образом, в среднесуглинистых по гранулометрическому составу почвах Москворецко-Окской равнины полночленный комплекс дождевых червей представлен только в наиболее старых лесах (возраст более 110 лет). Несмотря на присутствие быстроразлагаемого опада лиственных пород (липы, березы), на начальных стадиях сукцессии в этих лесах активно функционируют только две морфо-экологические группы дождевых червей: собственно почвенные и почвенно-подстилочные. При увеличении возраста лесных сообществ и доли медленно разлагаемого опада (ели обыкновенной) в горизонте подстилки сохраняются благоприятные условия для обитания подстилочных, почвенно-подстилочного и норных видов, которые питаются листовым опадом деревьев, кустарников и трав на поверхности почвы (Перель, 1979; Hoeffner et al., 2018; Huang et al., 2020).
1.3 Леса Северо-Западного Кавказа
Обнаружено 8 видов дождевых червей, принадлежащих к 4-м морфо-экологическим группам: подстилочные D.r. tenuis, D. octaedra, Dendrobaena attemsi Michaelsen, 1902; почвенно-подстилочный Eisenia fetida (Savigny, 1826), собственно почвенные Dendrobaena schmidti schmidti (Michaelsen, 1907), Dendrobaena tellermanica Perel, 1966, Aporrectodea jassyensis (Michaelsen, 1891) и норный Dendrobaena mariupolienis Wyssotzky, 1898.
В осиново-грабовых жимолостно-мелкотравных лесах (C1) обнаружены 7 видов дождевых червей: подстилочные D. octaedra, D. attemsi; почвенно-подстилочный E. fetida; собственно почвенные D. s. schmidti, D. tellermanica, A. jassyensis и норный D. mariupolienis (табл. 1). Биомасса собственно почвенных видов значимо выше, чем других групп (рис. 1).
В буково-пихтово-грабовых мелкотравных лесах (C2) обнаружены 5 видов дождевых червей: подстилочные D. octaedra, D. attemsi; собственно почвенные D. schmidi, A. jassyensis и норный D. mariupolienis.
В наиболее старых лесах (возраст более 400-х лет) пихтово-буковых мертвопокровных лесах (C3) обитает 7 видов дождевых червей: подстилочные D. octaedra, D. attemsi, D.r. tenuis; почвенно-подстилочный E. fetida; собственно почвенные D. s. schmidti, A. jassyensis и норный D. mariupolienis. Существенным отличием населения дождевых червей терминальной стадии сукцессии хвойно-широколиственных лесов Северо-Западного Кавказа является увеличение биомассы норных червей в 4-9 раз в сравнении с предыдущими стадиями хроноряда (Гераськина, 2018), различия статистически значимы (рис. 1, табл. 2).
В целом в хроносерии лесов Северо-Западного Кавказа уже на начальных этапах присутствовали 4 морфо-экологические группы дождевых червей, что связано с благоприятными свойствами лесных буроземов и присутствием смешанного опада, более благоприятного в трофическом и топическом отношении для дождевых червей (Sariyildiz, 2008; Sariyildiz, Küçük, 2008). Кислотность подстилки на всех стадиях сукцессии близка к оптимальным значениям (величина pH составляет от 5.1 до 6.0) и не лимитирует активность дождевых червей. Как и в лесах Москворецко-Окской равнины, на Северо-Западном Кавказе в ходе сукцессии происходило увеличение биомассы норных червей.
Таким образом, на примере трех типов лесных объектов исследования в разных регионах показано, что набор морфо-экологических групп дождевых червей определяется гранулометрическим составом почвы, качеством опада древесного яруса, подроста и кустарников. На начальных этапах послерубочного восстановления лесов состав морфо-экологических групп дождевых червей неполный (за исключение лесов Северо-Западного Кавказа). В ходе изменения сукцессионного статуса лесов происходит усложнение набора морфо-экологических групп дождевых червей, но не последовательная замена одних групп другими.
2. Влияние дождевых червей разных морфо-экологических групп на аккумуляцию углерода в лесных почвах и сопряженные почвенные параметры: запасы подстилки, содержание азота, С/N
Мощность подстилки регулируется активностью дождевых червей (Всеволодова-Перель и др., 1995; Suarez et al., 2006; Holdsworth et al., 2012; Huang et al., 2020 и др.). Корреляционный анализ показал, что во всех регионах мощность подгоризонта L отрицательно связана с биомассой дождевых червей, активных в горизонте подстилки. Наиболее высокие значения коэффициента детерминации (R2 = 0.65) получены для лесов Брянского Полесья, где запасы подгоризонта L не менее чем в два раза выше, в сравнении с лесами Москворецко-Окской равнины и Северо-Западного Кавказа (за исключением площадок старовозрастных буково-пихтовых мертвопокровных лесов) (рис. 2). Накопление подстилки в лесах Брянского Полесья соответствует очень низкой биомассе дождевых червей (рис. 1). Как показали наши исследования, биомасса и других групп беспозвоночных-сапрофагов здесь невелика. Запасы подстилки в лесах Москворецко-Окской равнины, вероятно, в более значительной степени регулируются деятельностью почвенно-подстилочного вида L. rubellus, поскольку его вклад в общую биомассу червей значимо выше в большей части лесных сообществ, чем вклад подстилочных и норных видов (рис. 1).
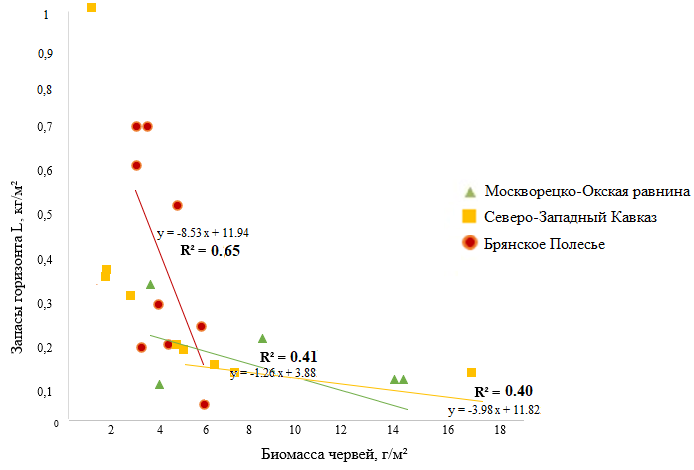
Рисунок 2. Зависимость мощности L-подгоризонта подстилки от биомассы подстилочных, почвенно-подстилочных и норных видов дождевых червей
В лесах Северо-Западного Кавказа из групп дождевых червей, регулирующих запасы подстилки, наибольший вклад вносят норные виды в связи с их значимо большей биомассой, чем видов подстилочной и почвенно-подстилочной групп (рис. 1).
Обнаружена значимая отрицательная корреляция между биомассой червей почвенно-подстилочных видов и соотношением С/N в подстилке и в гумусовом горизонте на примере лесов Москворецко-Окской равнины, где выявлена наибольшая биомасса L. rubellus (рис. 3а, 4а). Биомасса червей почвенно-подстилочного вида L. rubellus, напротив, положительно связана с содержанием азота в подстилке и гумусовом горизонте (рис. 3b, 4b). При этом направленность корреляции с содержанием органического углерода различается: в L-подгоризонте подстилки показана отрицательная корреляция содержания углерода с биомассой почвенно-подстилочных червей (рис. 3с), в гумусовом горизонте – напротив, положительная (рис. 4с). Поскольку почвенно-подстилочные черви активно питаются на поверхности почвы, что и может приводить к уменьшению содержания органического углерода в подстилке, но при этом относятся к первичным гумусообразователям (Перель, 1979), вероятнее всего, эффект аккумуляции органического вещества в ходе гумусообразования более выражен в гумусовом горизонте. Кроме того, в результате двигательной активности происходит перенос органического углерода из горизонта подстилки в гумусовый горизонт.
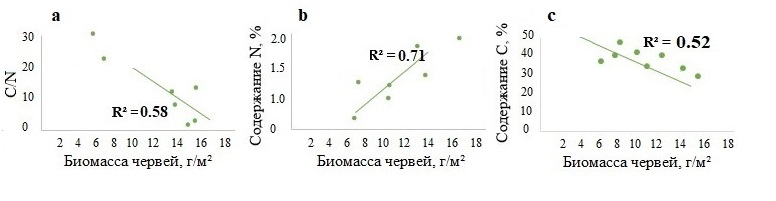
Рисунок 3. Зависимость показателя С/N (a), содержания азота (b) и углерода (c) в L-подгоризонте подстилки и биомассы почвенно-подстилочных дождевых червей в лесах Москворецко-Окской равнины
Значимые корреляции биомассы собственно почвенных видов выявлены только с показателем С/N в горизонте А (рис. 5), где черви этой группы наиболее активны при условии достаточной влажности. Значимые корреляционные взаимосвязи собственно почвенных видов с содержанием азота и углерода не установлены.
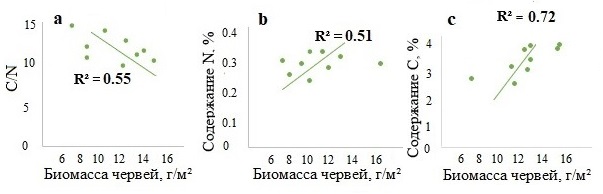
Рисунок 4. Зависимость показателя С/N (a), содержания азота (b) и углерода (c) в горизонте A от биомассы почвенно-подстилочных дождевых червей в лесах Москворецко-Окской равнины
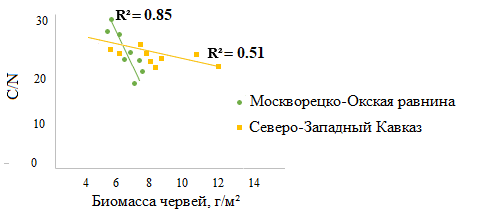
Рисунок 5. Зависимость показателя С/N в горизонте A от биомассы собственно почвенных дождевых червей в лесах Москворецко-Окской равнины и Северо-Западного Кавказа
Недостаток минеральных форм азота – один из важнейших лимитирующих факторов минерального питания растений, поскольку в почвах до 90% этого элемента находится в недоступной для растений форме (Mengel, 1996). Известно, что деятельность дождевых червей приводит к обогащению почвы доступными для растений формами азота. Копролиты дождевых червей обогащены мочевиной и ионами аммония. Пищеварительные ферменты червей активизируют деятельность нитрифицирующих и аммонифицирующих бактерий, следовательно, уменьшаются потери свободного азота, который закрепляется в форме соединений, аммонийный азот переходит в нитриты и нитраты (Козловская, 1976). Эксперименты с дождевыми червями Eisenia nordenskioldi, обитающими в темно-серой почве под широколиственными лесами, показали обогащение почвы подвижными формами аминного азота, доступного для поглощения корнями растений, свободными и связанными аминокислотами (Козловская и др., 1983; Стриганова и др., 1989). В опытах с почвенно-подстилочными и собственно почвенными дождевыми червями показано, что аммоний, содержащийся в копролитах дождевых червей, способен модифицировать нитрификацию в почве, вызывая долговременные кумулятивные эффекты, намного превосходящие свое прямое действие (Битюцкий и др., 2007). В экспериментах с норными червями показано, что содержание доступного азота в почве увеличивалось на 0.03 мг/кг на каждые 0.1 г биомассы дождевого червя (Andriuzzi et all., 2016). В природных экосистемах поток почвенного азота через популяции дождевых червей достигает в год десятков килограмм на гектар (Lee, 1985; Parmelee, Crossley, 1988), что необходимо для устойчивого функционирования наземных экосистем. Также обогащение почвы азотом происходит за счет гибели дождевых червей: ежегодно смертность которых составляет в среднем 60% от общей численности популяции (Lavelle et al., 1998). В почвах Центральной Европы после отмирания дождевых червей выход азота достигает 24 г/м2, что сопоставимо с ежегодной дозой минеральных азотных удобрений (100–200 кг N на 1 га). Биомасса дождевых червей, содержащая 65-75% белка, в почве быстро разлагается, но азот вымывается медленнее, поскольку связывается микроорганизмами (Lee, 1985; Makeschin, 1997; и др.).
Дождевые черви способствуют значительному снижению отношения С/N в почве, что связано с прямым и опосредованным влиянием дождевых червей на минерализацию и гумификацию органического вещества. Дождевые черви способствуют сужению отношения С/N в три раза по сравнению с опадом (Стриганова, 1968). Имеются экспериментальные доказательства значительного сужения C/N под влиянием разных морфо-экологических групп дождевых червей не только в лесных почвах. Для почвенно-подстилочных червей этот факт установлен в вермикомпостах (Talashilkar et al., 1999), для собственно почвенных червей — на сельскохозяйственных полях (Sandor, Schrader 2007; McDaniel et al. 2013).
Дождевых червей чаще относят к группе нитролеберантов – почвенных организмов, оказывающих сильное влияние на миграцию азота (Козловская, 1976; Жуков и др., 2000), в первую очередь за счет гумификации органического вещества в почве. Однако, дождевые черви как первичные разрушители подстилки и вторичные разрушители мертвых растительных остатков, оказывают влияние и на миграцию углерода в почвах, поэтому их можно отнести и к группе карболиберантов (минерализаторов). Согласно нашим и литературным данным, влияние разных морфо-экологических групп дождевых червей на содержание азота и показатель С/N однонаправленно в горизонтах их активности: содержание азота увеличивается, соотношение С/N уменьшается. Однако в отношении влияния дождевых червей на содержание углерода требуется дифференциальный функциональный подход. Последний глобальный метаанализ показывает, что присутствие не только подстилочных и норных групп, но и собственно почвенных дождевых червей приводит к снижению органического вещества в горизонте подстилки, при этом наиболее сильный эффект оказывают норные черви (Huang et al., 2020).
В нашем исследовании, кроме того, показан возможный значимый отрицательный эффект и группы почвенно-подстилочных червей (L. rubellus) на запасы подстилки и содержание органического углерода в ней. Известно, что этот вид часто приурочен к богатым почвам и высокому содержанию органического вещества (Жуков, 2004; Жуковская и др., 2005 и др.). Нами выявлены связи между биомассой почвенно-подстилочного L. rubellus и увеличением содержания углерода в гумусовом горизонте. Вероятно, это обусловлено высокой трофической активностью этих червей и хорошим качеством быстро разлагаемого опада (березы, липы, лещины). Между биомассой собственно почвенных видов и уровнем аккумуляции углерода значимых связей не установлено, но выявлена общая тенденция к снижению содержания органического углерода в гумусовом горизонте при увеличении биомассы этой группы видов. Собственно почвенные виды питаются почвенным перегноем (Перель, 1979; Жуков, 2004), в их копролитах по сравнению с почвой происходит уменьшение общей массы органического вещества и повышение зольности на 2-3% (Lavelle, Martin, 1992; Angst et al., 2017). Собственно почвенные виды не участвуют в активном перемещении подстилки и переносе органического углерода в нижележащие горизонты. Для получения убедительных результатов необходимы дополнительные полевые эксперименты в лесных почвах, которые запланированы нами в продолжение исследований по данной проблеме.
ВЫВОДЫ:
- В ходе изменения сукцессионного статуса лесов происходит усложнение видового состава и набора морфо-экологических групп дождевых червей, но не последовательная замена одних групп другими.
- Видовое богатство, разнообразие морфо-экологических групп и биомасса дождевых червей при сходном гранулометрическом составе почв определяется качеством опада: наиболее благоприятный для поддержания функционального разнообразия дождевых червей – смешанный опад лиственных и хвойных видов древесного полога, подроста и кустарников.
- Выявлены неоднозначные влияния дождевых червей разных морфо-экологических групп на аккумуляцию углерода в лесных почвах. Установлены отрицательные корреляционные зависимости между суммарной биомассой подстилочных, почвенно-подстилочных и норных видов червей и содержанием углерода подстилки. При этом биомасса почвенно-подстилочных видов положительно связана с содержанием углерода в гумусовом горизонте. Взаимосвязи содержания углерода почвы с собственно почвенными видами в проведенном исследовании не выявлены.
- Показатели, сопряженные с аккумуляцией углерода, – соотношение С/N и содержание азота однонаправленно коррелируют с биомассой дождевых червей в горизонтах их активности: при увеличении биомассы червей разных морфо-экологических групп соотношение С/N уменьшается, содержание азота повышается.
БЛАГОДАРНОСТИ
Исследование выполнено в рамках темы государственного задания ЦЭПЛ РАН № 0110-2018-0007, материал определен за счет средств гранта Российского научного фонда (проект № 16-17-10284).
СПИСОК ЛИТЕРАТУРЫ
Абатуров Б.Д. Почвообразующая роль животных в биосфере / Биосфера и почвы. М.: Наука, 1976. С. 53-69.
Березина О.Г. Структура населения коллембол (Hexapoda, Collembola) реликтового липового леса (Горная Шория, Кемеровская область) // Евразиатский энтомологический журнал. 2016. Т. 15. №. 6. С. 583-590.
Битюцкий Н.П., Соловьева А.Н., Лукина Е.И., Олейник А.С., Завгородняя Ю.А., Демин В.В., Бызов Б.А. Экскреты дождевых червей стимулятор минерализации соединений азота в почве // Почвоведение. 2007. № 4. С. 468-473.
Всеволодова-Перель Т.С. Дождевые черви фауны России. Кадастр и определитель. М.: Наука, 1997. 101 с.
Всеволодова-Перель Т.С., Грюнталь С.Ю., Кудряшева И.В., Надточий С.Э., Головач С.И., Матвеева А.А., Осипов В.В., Карпачевский Л.О., Растворова О.Г. Структура и функционирование почвенного населения дубрав Среднерусской лесостепи. М.: Наука, 1995. 152 с.
Гераськина А.П. Преобразования комплекса дождевых червей в ходе послерубочных сукцесcий в лесах Северо-Западного Кавказа // Вопросы лесной науки. 2018. № 1. С. 1-14.
Гиляров М.С. Методы почвенно-зоологических исследований. М.: Наука, 1975. 304 с.
Гончаров А.А. Структура трофических ниш в сообществах почвенных беспозвоночных (мезофауна) лесных экосистем / дис. на соиск. уч. степ. канд. биол. н. М.: ИПЭЭ, 2014. 177 с.
Жуков А.В. Дождевые черви как компонент биогеоценоза и их роль в зооиндикации // Ґрунтознавство. 2004. Т. 5 № 1-2. С. 44-57.
Жуков А.В. Экологическое разнообразие животного населения почв пойменных биогеоценозов р. Самара // Вісник Дніпропетровського університету. Біологія. Екологія. 2000. №. 7. С. 73-79.
Жуковская E.А., Кодолова О.П., Правдухина О.Ю., Варне А.Ж., Болотецкий Н.М. Исследование генетического разнообразия дождевого червя Lumbricus rubellus Hoff. (Oligochaeta, Lumbricidae) // Известия Российской академии наук. Серия биологическая. 2005. № 5. С. 625-627.
Классификация и диагностика почв России / Сост. Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Смоленск: Ойкумена, 2004. 342 с.
Козловская Л.С. Роль почвенных беспозвоночных в трансформации органического вещества болотных почв. Л., 1976. 211 с.
Козловская Л.С., Арчегова И.Б., Ракова H.Н. Биохимическое воздействие почвенных беспозвоночных на растительные остатки // Болотные биогеоценозы и их изменения в результате антропогенного воздействия. Л.: Наука, 1983. С. 24-26.
Кузнецова А.И., Лукина Н.В., Тихонова Е.В., Горнов А.В., Горнова М.В., Смирнов В.Э., Гераськина А.П., Шевченко Н.Е., Тебенькова Д. Н., Чумаченко С.И. Аккумуляция углерода в песчаных и суглинистых почвах равнинных хвойно-широколиственных лесов в ходе послерубочных восстановительных сукцессий // Почвоведение. 2019. № 7. С. 803-816.
Лукина Н.В., Тихонова Е.В., Шевченко Н.Е., Горнов А.В., Кузнецова А.И., … & Шанин В.Н. Аккумуляция углерода в лесных почвах и сукцессионный статус лесов / под ред. член-корр. Н.В. Лукиной. М.: КМК, 2018. 232 с.
Методические указания по количественному определению объема поглощения парниковых газов // Распоряжение Минприроды России от 30.06.2017 № 20-р. 108 с.
Национальный атлас почв Российской Федерации / С.А. Шоба, И.О. Алябина, И.С. Урусевская, О.В. Чернова. Москва: Астрель, 2011. 632 с.
Перель Т.С. Распространение и закономерности распределения дождевых червей фауны СССР. М.: Наука, 1979. 272 с.
Стриганова Б.Р. Исследование роли мокриц и дождевых червей в процессах гумификации разлагающейся древесины // Почвоведение. 1968. №. 8. С. 85-90.
Стриганова Б.Р. Питание почвенных сапрофагов. М.: Наука, 1980. 243 с.
Стриганова Б.Р. Сукцессии животного населения почвы в процессе первичного почвообразования // Ранняя колонизация суши. М.: ПИН РАН. 2012. С. 177-195.
Стриганова Б.Р., Козловская Л.С., Чернобровкина Н.П., Кудряшева И.В. Пищевая активность дождевых червей и содержание аминокислот в тёмно-серой лесной почве. Почвоведение. 1989. № 5. С. 44-51.
Шевченко Н.Е., Кузнецова А.И., Тебенькова Д.Н., Смирнов В.Э., Гераськина А.П., Горнов А.В., Тихонова Е.В., Лукина Н.В. Сукцессионная динамика растительности и запасы почвенного углерода в хвойно-широколиственных лесах северо-западного Кавказa // Лесоведение. 2019. № 3. С. 163-176.
Alban D.H., Berry E.C. Effects of earthworm invasion on morphology, carbon, and nitrogen of a forest soil // Appl. Soil Ecol. 1994. Vol. 1. P. 243-249.
Andriuzzi W.S., Schmidt O., Brussaard L., Faber J.H., Bolger T. Earthworm functional traits and interspecific interactions affect plant nitrogen acquisition and primary production // Applied Soil Ecology. 2016. Vol. 104. P. 148-156.
Angst S., Mueller C.W., Cajthaml T., Angst G., Lhotakova Z., Bartuska M., Spaldonova A., Frouz J. Stabilization of soil organic matter by earthworms is connected with physical protection rather than with chemical changes of organic matter // Geoderma. 2017. Vol. 289. P. 29-35.
Begon M., Harper J.L., Townsend C.R. Ecology: Individuals, Populations and Communities. Oxford: Blackwell Scientifi c Publications, 1986. 1068 p.
Bohlen P.J., Pelletier D.M., Groffman P.M., Fahey T.J., Fisk M.C. Influence of earthworm invasion on redistribution and retention of soil carbon and nitrogen in northern temperate forests // Ecosystems. 2004. Vol. 7. P. 13-27.
Burtelow A.E., Bohlen P.J., Groffman P.M. Influence of exotic earthworm invasion on soil organic matter, microbial biomass and denitrification potential in forest soils of the northeastern United States // Appl. Soil Ecol. 1998. Vol. 9. P. 197-202.
Curry J.P. Factors affecting the abundance of earthworms in soils. In: Edwards C.A. (Ed.). Earthworm Ecology. 2nd ed. CRC Press, Boca Raton, FL, 1994. P. 91-113.
Frouz J., Liveckova M., Albrechtoa J., Chronakova A., Cajthaml T., … & Simackova H. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites // Forest Ecology and Management. 2013. Vol. 309. P. 87-95.
Gleixner G. Soil organic matter dynamics: a biological perspective derived from the use of compound-specific isotopes studies // Ecological Research. 2013. Vol. 28. No. 5. P. 683-695.
Hirth J.R., Li G.D., Chan K.Y., Cullis B.R. Long-term effects of lime on earthworm abundance and biomass in an acidic soil on the south-western slopes of New South Wales, Australia // Applied Soil Ecology. 2009. Vol. 43. P. 106-114.
Hoeffner K., Monard C., Santonja M., Cluzeau D. Feeding behaviour of epi-anecic earthworm species and their impacts on soil microbial communities // Soil Biology and Biochemistry. 2018. Vol. 125. P. 1-9.
Holdsworth A.R., Frelich L.E., Reich P.B. Leaf litter disappearance in earthworm-invaded northern hardwood forests: role of tree species and the chemistry and diversity of litter // Ecosystems. 2012. Vol. 15. No. 6. P. 913-926.
Huang W., Gonzalez G., Zou X. Earthworm abundance and functional group diversity regulate plant litter decay and soil organic carbon level: A global meta-analysis //Applied Soil Ecology. 2020. Vol. 150. P. 1-15.
Lavelle P., Martin A. Small-scale and large-scale effects of endogeic earthworms on soil organic matter dynamics in soils of the humid tropics // Soil Biology and Biochemistry. 1992. Vol. 24. No. 12. P. 1491-1498.
Lavelle P., Pashanasi B., Charpentier F., Gilot C., Rossi J.-P., …& Bernier N. Large- scale effects of earthworms on soil oranic matter and nutrient dynamics / Edwards C.A. (Ed.). Earthworm ecology. St. Lucie Press, 1998. P.103-122.
Lee K.E. Earthworms. Their ecology and relationships with soils and land use. Academic Press, 1985. 411 p.
Makeschin F. Earthworms (Lumbricidae: Oligochaeta): Important promoters of soil development and soil fertility / G. Benckiser (Ed.). Fauna in soil ecosystems. Recycling processes, nutrient fluxes and agricultural production. Florida: CRC Press, 1997. P. 173-223.
McDaniel J.P., Stromberger M.E., Barbarick K.A., Cranshaw W. Survival of Aporrectodea caliginosa and its effects on nutrient availability in biosolids amended soil // Applied soil ecology. 2013. Vol. 71. P. 1-6.
Mengel K. Turnover of organic nitrogen in soils and its availability to crops // Plant and Soil. 1996. Vol. 181. P. 83-96.
Moore J.-D., Ouimet R., Bohlen P.J. Effects of liming on survival and reproduction of two potentially invasive earthworm species in nothern forest Podzol // Soil Biology and Biochemistry. 2013. Vol. 64. P. 174-180.
Novara A., Rühl J., La Mantia, T., Gristina L., La Bella, S., Tuttolomondo T. Litter contribution to soil organic carbon in the processes of agriculture abandon // Solid Earth. 2015. Vol. 6. No. 2. P. 425-432.
Parmelee R.W., Crossley Jr. Earthworm production and role in the nitrogen cycle of a no-tillage agroecosystem on the Georgia piedmont // Pedobiologia. 1988. Vol. 32. P. 353-361.
Pulleman M.M., Six J., Uyl A., Marinissen J.C.Y., Jongmans A.G. Earthworms and management affect organic matter incorporation and microaggregate formation in agricultural soils // Appl. Soil Ecol. 2005. Vol. 29. Р. 1-15.
Sandor M., Schrader S. Earthworms affect mineralization of different organic amendments in a microcosm study // Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Agriculture. 2007. Vol. 63. P. 442-447.
Sariyildiz T. Effects of tree canopy on litter decomposition rates of Abies nordmanniana, Picea orientalis and Pinus sylvestris // Scandinavian journal of forest research. 2008. Vol. 23. No. 4. P. 330-338.
Sariyildiz T., Küçük M. Litter mass loss rates in deciduous and coniferous trees in Artvin, northeast Turkey: Relationships with litter quality, microclimate, and soil characteristics // Turkish journal of Agriculture and Forestry. 2008. Vol. 32. No. 6. P. 547-559.
Suarez E.R., Fahey T.J., Yavitt J.B., Groffman P.M., Bohlen P.J. Patterns of litter disappearance in a northern hardwood forest invaded by exotic earthworms // Ecological Applications. 2006. Vol. 16. No. 1. P. 154-165.
Talashilkar S.C., Bhangarath P.P., Mehta V.B. Changes in chemical properties during composting of organic residues as influenced by earthworm activity // Journal of the Indian Society of Soil Science. 1999. Vol. 47. P. 50-53.
World Reference Base for Soil Resources. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports. IUSS Working Group. Rome: FAO, 2015. 203 р.
Рецензент: к.б.н., доцент, в.н.с. Зенкова И.В.





