- DOI 10.31509/2658-607x-202363-132
- УДК 630*114.351:58.072:57.042:502.055 (470.21)
Сравнительная оценка скорости разложения растительного опада в еловых и сосновых лесах на северном пределе распространения
![]()
© 2023 г. Е. А. Иванова1,2*, М. А. Данилова2, В. Э. Смирнов2, В. В. Ершов1
1Институт проблем промышленной экологии Севера, ФИЦ КНЦ РАН
Россия, 184209, Мурманская обл., Апатиты, ул. Академгородок, 14а
2Центр по проблемам экологии и продуктивности лесов Российской академии наук
Россия, 117997, Москва, ул. Профсоюзная, 84/32, стр. 14
*E-mail: ea.ivanova@ksc.ru
Поступила в редакцию 16.08.2023
После рецензирования: 19.09.2023
Принята к печати: 20.09.2023
Проведена сравнительная оценка процессов разложения растительных остатков (хвоя сосны, хвоя ели, листья бореальных кустарничков, слоевища мхов) на начальных этапах в сосняках лишайниково-кустарничковых и ельниках кустарничково-зеленомошных, формирующихся в естественных условиях на северном пределе распространения. Изучались особенности исходного состава опада, темпы разложения и изменения химического состава растительных остатков в процессе деструкции, обусловленные формацией леса. Более высокое исходное содержание Сорг в тканях растений сосновых лесов связано с благоприятными условиями освещенности под пологом леса, тогда как высокое содержание Mn в тканях растений напочвенного покрова в ельниках обусловлено непосредственным влиянием богатого этим элементом питания опада хвои ели. Результаты исследования наглядно продемонстрировали, что формация леса оказывает значительное влияние как на исходное качество опада одних и тех же видов растений, так и на скорость разложения: хвоя ели и листья брусники с более высоким содержанием элементов питания (Mg, Mn, P) и узкими соотношениями элементов (С:N, C:P) в еловом лесу характеризовались и более активными процессами разложения. Однако опад зеленых мхов, несмотря на более высокое качество в ельниках, разлагался активнее в сосновых лесах, что может быть связано с большим количеством осадков в сосновых лесах. Таким образом, на различия в скорости разложения растительных остатков влияет сочетанием качества растительного материала, температурного режима и количества осадков, связанных с формацией леса.
Ключевые слова: формация леса, разложение опада, растительные остатки, качество опада
Опад древесных растений и растений напочвенного покрова — листья, хвоя, почки, побеги, плоды, корни и др., отмирающие в течение определенного времени, является одним из важнейших компонентов, влияющих на формирование биогеохимических циклов лесных экосистем. Растительный опад является источником органического углерода и элементов минерального питания, которые становятся доступными для биоты в ходе его разложения и минерализации.
Скорость разложения опада и поступления углерода в почву во многом зависят от гидротермических условий почв (Кузнецов, 2010; Кузнецов, Осипов, 2011), активности и состава почвенной биоты (Воробьева, Наумова, 2009; Högberg et al., 2017), фракционного состава поступающего опада (Бобкова, 2000; Fang et al., 2015), климатических условий (температуры воздуха и количества осадков) (Pausas, 1997; Portillo-Estrada et al., 2016). По современным представлениям ведущим фактором регионального масштаба считается климат, локального — качество опада, тогда как деятельность организмов-деструкторов регулируется климатом и качеством опада (Bradford et al., 2016), определяемого концентрациями элементов питания и вторичных метаболитов. Так, на ранних стадиях разложения наибольшие эффекты оказывают N, P и водорастворимые органические соединения, на более поздних основным детерминантом динамики разложения является лигнин (Berg, 2000; Wardle et al., 2003; Zhang et al., 2008; Rahman et al., 2013; Ларионова и др., 2017). Величина стехиометрических отношений элементов и веществ (C:N, лигнин:N) в разлагающемся опаде также связана с качеством растительного материала: чем у́же отношения, тем выше скорость деструкции (Berg, McClaugherty, 2008; Rahman et al., 2013; Tu et al., 2014; Lukina et al., 2017). Исходный химический состав опада, определяющий его качество, а соответственно и процесс его разложения, проявляет видовые особенности. У четырех различных видов сосны (Pinus pinea, P. laricio, P. sylvestris и P. nigra) скорость разложения регулировалась исходным содержанием элементов питания (N, K, Mn) (De Marco et al., 2007). Хвоя сосны (P. contorta) по сравнению с еловой хвоей (Picea engelmannii) содержала больше C и N и меньше лигнина, что говорит о более высоком качестве растительного материала, однако потери углерода были выше для ели (Leonard et al., 2020). В другом исследовании показано, что опад хвои ели с более высоким содержанием элементов питания и относительно узкими отношениями C:N и лигнин:N разлагался заметно быстрее, чем опад хвои сосны, тогда как опад листьев березы повислой (Betula pendula) в сосновых лесах, с более низким соотношением N:P, разлагался быстрее по сравнению с опадом березы пушистой (B. pubescens) в ельниках (Иванова и др., 2019).
Древесные растения способны формировать типы леса со своими особенными условиями местообитания и плодородием почв, в том числе через опад, химический состав которого влияет на микробную активность и состав почв (Рахлеева и др., 2011; Aponte et al., 2013; Chavez-Vergara et al., 2014; Колмогорова, Уфимцев, 2018; Помогайбин, Помогайбин, 2018; Цандекова, 2018). Полог древостоя в зависимости от видового состава, строения и сомкнутости крон изменяет состав атмосферных выпадений, влияет на уровень освещенности (Лукина и др., 2008; Кищенко, 2019). Лесные экосистемы на территории Мурманской области представляют собой стадии сукцессий, в ходе которых происходит смена древесных пород (сосна, ель, береза), которые на почвообразующих породах сходного состава создают разные условия среды в экосистемах. Так, ель в большей степени по сравнению с сосной подкисляет проходящие через кроны осадки, препятствует проникновению их под кроны деревьев, создавая более ярко выраженную мозаичность. Ель за счет большей сомкнутости крон препятствует проникновению солнечного света под полог леса. В еловых лесах плодородие почв значительно выше, чем в сосновых, формирующихся на тех же почвообразующих породах и позициях в рельефе (Лукина и др., 2002, 2006, 2008, 2010; Цветков, 2004; Орлова и др., 2011). Это может определять и различия в составе опада одних и тех же видов растений в различных типах леса, и оказывать влияние на скорость их разложения.
Цель данной работы — оценить влияние формации леса на исходный состав и скорость разложения опада в доминирующих в северотаежной подзоне еловых и сосновых лесах.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились на площадках постоянного наблюдения (ППН) в Мурманской области в сосняках лишайниково-кустарничковых (10С) на подзолах иллювиально-железистых (Rustic podzols) в 1997–1999 гг. и в ельниках зеленомошно-кустарничковых (8Е2Б) на подзолах иллювиально-гумусовых (Carbic podzols) в автоморфных условиях в 1996–1998 гг. Более подробные описания объектов исследования даны в ранних работах (Лукина, Никонов, 1996, 1998; Lukina et al., 2017; Иванова и др., 2019). Основными почвообразующими породами на территории исследования являются ледниковые отложения — моренные и водно-ледниковые, по гранулометрическому составу — песчаные и супесчаные (Белов, Барановская, 1969; Переверзев, 2004). В процессе сукцессии хвойных лесов при смене сосняков ельниками наблюдаются закономерности изменения химического состава почв: в еловых лесах увеличивается содержание доступных соединений биофильных элементов во всем почвенном профиле, при этом в органогенных горизонтах снижается отношение C:N и наблюдается накопление гумуса. В сосновых лесах органогенные горизонты подкроновых пространств характеризуются более высокой кислотностью, чем межкроновых, тогда как в еловых лесах наблюдается обратная картина. В минеральных горизонтах почв наблюдается увеличение содержания кислотных компонентов, в частности в иллювиальных горизонтах отмечено увеличение содержания фульвокислот на фоне снижения содержания гуминовых кислот (Орлова и др., 2011).
Эксперименты по изучению процессов разложения опада на ППН в разных типах леса проводились с соблюдением единообразия методов и статистической обработки данных. Образцы активной фракции опада (листья и хвоя) доминирующих видов сосудистых растений (P. sylvestris, P. abies, Vaccinium vitis-idaea, V. myrtillus и Empetrum hermaphroditum) и образцы стареющей бурой части мхов (Pleurozium schreberi) отбирались в сентябре 1997 г. на мониторинговом стационаре в сосняках, в октябре 1996 г. — в ельниках. Растительный материал (10 г сухого вещества) для разложения закладывали в пакеты из синтетического материала с размером пор 30 мкм, которые размещали на поверхности почвы (в подгоризонт L подстилки) в подкроновых и межкроновых пространствах на мониторинговых площадках. Пробы отбирались ежегодно в октябре через 1 и 2 года после начала каждого эксперимента. Всего за два периода отбора было отобрано 32 пакета в сосновых лесах и 49 — в еловых.
Перед проведением химического анализа растительный материал измельчали и подвергали мокрому озолению концентрированной HNO3. Концентрации металлов (Ca, Mg, K, Mn) определяли методом атомно-абсорбционной спектрометрии на приборе Aanalyst 800. Общее содержание азота определялось методом Къельдаля, органического углерода (Cорг) — методом Тюрина, фосфора — методом колориметрии (Воробьева, 1998). Содержание лигнина определяли путем обработки пробы 72%-ной H2SO4 после предварительного кипячения в растворе цетилтриметиламмония бромида в 0.5-молярном растворе H2SO4 (Rowland, Roberts, 1994).
Расчеты проводились на абсолютно сухой вес. Потери массы рассчитывались как разница между массой образцов до закладки и через 1 или 2 года и выражались в процентах. Коэффициент обогащения, демонстрирующий изменение состава растительного материала в процессе разложения, вычислялся для каждого элемента как отношение концентрации после первого или второго года разложения к исходной. Потери элементов питания и лигнина с учетом темпов потери массы выражались в процентах и рассчитывались как разница произведений концентрации компонента на массу образца до начала эксперимента и через 1 или 2 года соответственно. Качество опада характеризовали на основе содержания лигнина (вторичные метаболиты), элементов питания (N, Ca, Mg, K, P, Mn), и стехиометрических соотношений C:N, C:P, лигнин:N, N:P.
Статистический анализ данных проводился различными методами. Оценка влияния формации лесов (еловые и сосновые) на исходный химический состав растительного материала проводилась для отдельных видов растений с применением V-критерия (Husson et al., 2017) в среде статистического программирования R (R Core Team …, 2017). При этом матрица данных по химическому составу опада была дополнена имеющимися данными по составу живых растений: хвои сосны и ели последних лет жизни, многолетних листьев брусники, вороники, многолетних и отмерших побегов зеленых мхов P. schreberi и Hylocomium splendens. Сравнение параметров разложения (потери массы и элементов, коэффициент обогащения) в сосновых и еловых лесах проводилось для каждой фракции опада (хвоя сосны и ели, листья брусники, вороники, мхи) с помощью U-теста Манна-Уитни в программе Statistica. Возможное влияние внутри- и межбиогеоценотической изменчивости температур почвы и воздуха в еловых и сосновых лесах проводилось с использованием данных, полученных в период 2015–2021 гг. с помощью температурных логгеров под кронами деревьев, в межкроновых пространствах и на деревьях.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Исходный состав растительного материала — неразложившегося опада и еще живых растений (их многолетних частей) — характеризует качество растительного материала для последующего разложения его организмами-деструкторами.
Сравнение состава отдельных фракций опада, а также живого растительного материала соответствующих видов растений показало, что хвоя ели последних лет жизни богаче элементами питания Ca, Mg, Mn, P и характеризуется более узкими соотношениями C:P и N:P (p<0.05), что подтверждает ранее полученные данные (Сухарева, Лукина, 2014), тогда как хвоя сосны характеризуется более высоким содержанием C и N (табл. 1), что согласуется с результатами других авторов (Leonard et al., 2020). При этом следует заметить, что, в отличие от ранее полученных результатов, концентрации К в данном исследовании оказались выше в многолетней хвое ели. Более высокое содержание азота в хвое сосны последних лет жизни (7-8 лет) по сравнению с 11-13-летней хвоей ели объяснялось ранее (Сухарева, Лукина, 2014) различиями в возрасте: N, как и K и P, относится к подвижным элементам питания (Helmisaari, 1990; Rautio, 1998), то есть происходит транслокация его соединений в более молодые ткани и обеднение более старой хвои. Содержание других подвижных элементов питания, которые относятся к элементам минерального питания — K, P, и Mg (элементы средней подвижности), напротив, оказалось выше в хвое ели. Содержание малоподвижных элементов минерального питания, к которым относятся Ca и Mn, как и ожидалось, значительно выше в стареющей хвое ели.
Таблица 1. Химический состав живых и отмерших многолетних частей растений в северотаежных сосновых и еловых лесах
| Параметр | Среднее | Стандартное отклонение | Общее среднее | Общее стандартное отклонение | р | n | ||||
| Е-БГЦ | С-БГЦ | Е-БГЦ | С-БГЦ | Е-БГЦ | С-БГЦ | |||||
| Хвоя ели и сосны последних лет жизни* | ||||||||||
| Ca | мг/кг | 13 878 | 4077 | 3904 | 941 | 11 321 | 5497 | 0 | 51 | 18 |
| Mg | 674 | 495 | 249 | 112 | 628 | 235 | 0.01 | 51 | 18 | |
| K | 3527 | 3212 | 604 | 340 | 3445 | 563 | 0.04 | 51 | 18 | |
| Mn | 2149 | 977 | 923 | 295 | 1844 | 958 | 0 | 51 | 18 | |
| P | 1139 | 928 | 243 | 104 | 1084 | 234 | 0.001 | 51 | 18 | |
| N | 8805 | 10 162 | 1248 | 1579 | 9159 | 1459 | 0.001 | 51 | 18 | |
| Cорг | % | 52 | 56 | 4 | 6 | 53 | 5 | 0.01 | 51 | 18 |
| C:N | 61 | 56 | 10 | 8 | 59 | 10 | 0.07 | 51 | 18 | |
| C:P | 478 | 607 | 97 | 96 | 511 | 112 | 0 | 51 | 18 | |
| N:P | 8 | 11 | 2 | 2 | 9 | 2 | 0 | 51 | 18 | |
| Брусника — многолетние листья (живые части) | ||||||||||
| Ca | мг/кг | 6535 | 6878 | 1420 | 1063 | 6699 | 1260 | 0.37 | 23 | 21 |
| Mg | 1291 | 1089 | 306 | 127 | 1195 | 257 | 0.01 | 23 | 21 | |
| K | 3917 | 3629 | 482 | 507 | 3780 | 509 | 0.06 | 23 | 21 | |
| Mn | 2091 | 1704 | 371 | 426 | 1907 | 439 | 0.004 | 23 | 21 | |
| P | 952 | 725 | 204 | 81 | 844 | 194 | 0.0001 | 23 | 21 | |
| N | 8832 | 8397 | 1716 | 2275 | 8625 | 1991 | 0.47 | 23 | 21 | |
| Cорг | % | 51 | 55 | 3 | 3 | 53 | 4 | 0.001 | 23 | 21 |
| C:N | 60 | 69 | 10 | 17 | 64 | 14 | 0.03 | 23 | 21 | |
| C:P | 556 | 760 | 106 | 85 | 653 | 140 | 0 | 23 | 21 | |
| N:P | 9 | 12 | 1 | 3 | 10 | 2 | 0.002 | 23 | 21 | |
| Вороника — многолетние листья (живые части) | ||||||||||
| Ca | мг/кг | 8167 | 8755 | 1285 | 1455 | 8342 | 1345 | 0.22 | 26 | 11 |
| Mg | 2381 | 2710 | 273 | 300 | 2479 | 317 | 0.004 | 26 | 11 | |
| K | 4571 | 3823 | 1279 | 593 | 4349 | 1163 | 0.07 | 26 | 11 | |
| Mn | 1016 | 995 | 464 | 187 | 1009 | 399 | 0.88 | 26 | 11 | |
| P | 959 | 1038 | 180 | 113 | 982 | 166 | 0.19 | 26 | 11 | |
| N | 9610 | 10 895 | 2388 | 2900 | 9992 | 2579 | 0.17 | 26 | 11 | |
| Cорг | % | 57 | 58 | 3 | 1 | 57 | 3 | 0.64 | 26 | 11 |
| C:N | 63 | 56 | 16 | 15 | 61 | 16 | 0.23 | 26 | 11 | |
| C:P | 613 | 561 | 109 | 63 | 597 | 100 | 0.14 | 26 | 11 | |
| N:P | 10 | 11 | 2 | 3 | 10 | 2 | 0.66 | 26 | 11 | |
| Вороника — многолетние листья (отмершие части) | ||||||||||
| Ca | мг/кг | 8132 | 8453 | 605 | 847 | 8315 | 744 | 0.43 | 6 | 8 |
| Mg | 1950 | 1774 | 332 | 86 | 1850 | 234 | 0.16 | 6 | 8 | |
| K | 3106 | 2034 | 1296 | 293 | 2493 | 998 | 0.05 | 6 | 8 | |
| Mn | 1212 | 678 | 293 | 49 | 907 | 331 | 0.003 | 6 | 8 | |
| P | 720 | 1020 | 175 | 22 | 891 | 189 | 0.003 | 6 | 8 | |
| N | 7577 | 9204 | 2685 | 825 | 8507 | 1959 | 0.12 | 6 | 8 | |
| Cорг | % | 57 | 62 | 5 | 5 | 60 | 5 | 0.08 | 6 | 8 |
| C:N | 81 | 69 | 19 | 10 | 74 | 15 | 0.14 | 6 | 8 | |
| C:P | 825 | 613 | 161 | 49 | 704 | 152 | 0.01 | 6 | 8 | |
| N:P | 10 | 9 | 1 | 1 | 10 | 1 | 0.04 | 6 | 8 | |
| Мхи — живые части | ||||||||||
| Ca | мг/кг | 2519 | 2315 | 543 | 292 | 2481 | 508 | 0.38 | 26 | 6 |
| Mg | 974 | 743 | 502 | 80 | 931 | 461 | 0.27 | 26 | 6 | |
| K | 5160 | 4280 | 1136 | 776 | 4995 | 1122 | 0.08 | 26 | 6 | |
| Mn | 558 | 483 | 266 | 128 | 544 | 246 | 0.50 | 26 | 6 | |
| P | 1168 | 852 | 268 | 121 | 1109 | 275 | 0.01 | 26 | 6 | |
| N | 7338 | 5784 | 2013 | 728 | 7046 | 1932 | 0.08 | 26 | 6 | |
| Cорг | % | 45 | 51 | 2 | 4 | 46 | 4 | 0.0001 | 26 | 6 |
| C:N | 65 | 91 | 18 | 18 | 70 | 20 | 0.01 | 26 | 6 | |
| C:P | 400 | 616 | 89 | 120 | 440 | 127 | 0.0002 | 26 | 6 | |
| N:P | 6 | 7 | 1 | 1 | 6 | 1 | 0.33 | 26 | 6 | |
| Мхи — отмершие части | ||||||||||
| Ca | мг/кг | 4288 | 4456 | 1129 | 1095 | 4384 | 1069 | 0.77 | 6 | 8 |
| Mg | 668 | 1884 | 139 | 788 | 1363 | 855 | 0.01 | 6 | 8 | |
| K | 3434 | 2645 | 855 | 887 | 2983 | 932 | 0.12 | 6 | 8 | |
| Mn | 922 | 615 | 302 | 135 | 747 | 264 | 0.03 | 6 | 8 | |
| P | 952 | 898 | 135 | 122 | 922 | 126 | 0.43 | 6 | 8 | |
| N | 6656 | 8600 | 497 | 2602 | 7767 | 2177 | 0.10 | 6 | 8 | |
| Cорг | % | 45 | 51 | 4 | 8 | 49 | 7 | 0.14 | 6 | 8 |
| C:N | 69 | 62 | 8 | 10 | 65 | 9 | 0.17 | 6 | 8 | |
| C:P | 485 | 566 | 82 | 39 | 531 | 72 | 0.04 | 6 | 8 | |
| N:P | 7 | 9 | 2 | 2 | 8 | 2 | 0.04 | 6 | 8 | |
Примечание: Е-БГЦ — еловый биогеоценоз; С-БГЦ — сосновый биогеоценоз; * — в Е-БГЦ представлены данные по составу хвои ели, в С-БГЦ — хвои сосны; р — вероятность ошибки 1-го рода при расчете V-критерия
Многолетние живые листья брусники в ельниках, в отличие от листьев в сосняках, характеризуются достоверно более узкими соотношениями элементов (C:N, C:P, N:P) и более высоким содержанием элементов питания (Mg, Mn, P), что согласуется с данными по составу листьев текущего года (Исаева, Сухарева, 2013). Однако содержание углерода выше в многолетних листьях брусники в сосновых лесах (p<0.05).
Опад и материал многолетних живых органов вороники и мхов проявили отчетливые различия в химическом составе в зависимости от типа леса только в отношении Mn и С: содержание марганца выше в еловых лесах, а углерода — в сосновых. Примечательно, что живая стареющая хвоя сосны также содержит больше углерода, чем еловая, которая оказалась богата марганцем. Известно, что ассимилирующие органы сосны обыкновенной показывают более высокую интенсивность процессов фотосинтеза по сравнению с еловыми (Тужилкина, 1984; Суворова, 2006; Молчанов, 2020; Yang et al., 2020). Более высокое содержание углерода в растительных тканях кустарничков и мхов в сосновых лесах может быть связано с более высокой интенсивностью фотосинтеза растений под пологом сосновых деревьев: густые низкоопущенные кроны ели перехватывают солнечную радиацию гораздо более эффективно, чем высоко поднятые ажурные кроны сосны обыкновенной (Кищенко, 2019). Соответственно, в условиях лучшей освещенности при меньшей сомкнутости крон растения напочвенного покрова эффективнее продуцируют органическое вещество и накапливают больше углерода в тканях (Аткина, 2000; Зубкова и др., 2022). В еловых лесах повышенное содержание Mn в растениях напочвенного покрова может быть связано с влиянием опада хвои ели, богатого этим элементом питания, на его содержание в почвах.
Содержание остальных элементов питания не проявляло четких зависимостей от типа леса. В отмерших листьях вороники содержание K также выше в ельниках, но содержание P выше в сосняках. Согласно данным, полученным ранее на этих объектах исследования, листья вороники текущего года характеризуются более высоким содержанием K, Mn и Р в сосняках кустарничково-лишайниковых, по сравнению с ельниками кустарничково-зеленомошными (Исаева, Сухарева, 2013). Соотношения элементов в многолетних отмерших листьях вороники также проявляли различия: соотношение C:P было ýже в ельниках, N:P — в сосняках. Зеленые мхи (живые многолетние части) в ельниках богаче фосфором, в сосняках — углеродом, что согласуется с литературными данными по составу зеленых мхов (Сухарева, 2018). При этом величина соотношений C:N и C:P меньше в ельниках. Отмершие части мхов характеризовались различиями в содержании других элементов: Mg в мхах больше в сосняках, Mn — в ельниках, соотношения C:P и N:P ýже в ельниках.
Таким образом, содержание элементов питания и углерода в стареющих и отмерших листьях кустарничков, так же как и в тканях мхов, различается в зависимости от типа леса, что наиболее отчетливо проявляется в отношении Mn и Cорг: в еловых лесах растительный материал кустарничков и мхов обогащен марганцем, в сосновых — углеродом. Это можно объяснить различиями в условиях произрастания, формируемыми доминирующими древесными растениями: плодородие и влажность почв еловых лесов выше, чем сосновых, в тех же климатических условиях и на тех же почвообразующих породах (Лукина и др., 2008). Однако в сосновых лесах за счет ажурности крон создаются более благоприятные условия освещенности для активного фотосинтеза растений напочвенного покрова и, соответственно, более активного накопления углерода. Также вероятна непосредственная связь химического состава растений напочвенного покрова с составом опада ассимилирующих органов древесных растений-эдификаторов: на повышенное содержание Mn в растениях напочвенного покрова влияет его поступление в почву с опадом хвои ели, богатым марганцем.
Скорость разложения и изменения химического состава растительных остатков в процессе деструкции
Различия в скорости потерь массы могут быть обусловлены исходным качеством и условиями внешней среды. По скорости потерь массы после первого года разложения опад изучаемых видов распределяется в ряду: брусника > хвоя ели > вороника > мхи; после второго года: хвоя ели > вороника > брусника > мхи. В сосновых биогеоценозах ряды скорости разложения выглядят следующим образом: после первого года: вороника > брусника > хвоя сосны > мхи; после второго года: хвоя сосны > брусника > вороника > мхи (табл. 2). Как в более раннем исследовании, проведенном в еловых лесах, при относительно высокой исходной величине соотношения C:N в опаде мхов как в ельниках, так и в сосняках, скорость потери их массы была ниже, чем у остальных видов (Lukina et al., 2017). В других работах также показано, что моховой опад разлагается медленно (Wardle et al., 2003; Cornelissen et al., 2007; Hilli, 2013), что может быть обусловлено высоким содержанием неидентифицированных фенольных соединений в клеточных стенках мохообразных и широким соотношением C:N в растительных остатках (Ligrone et al., 2008; Lukina et al., 2017).
Таблица 2. Потери массы, элементов питания и лигнина (в %), стехиометрические соотношения элементов при разложении активных фракций растительного опада в северотаежных сосновых и еловых лесах
| Параметр | Е-БГЦ | С-БГЦ | p | n | |||
| Среднее | Стандартная ошибка | Среднее | Стандартная ошибка | Е-БГЦ | С-БГЦ | ||
| 1 год | |||||||
| Хвоя* | |||||||
| Масса | 20 | 1 | 16 | 1 | 0.01 | 5 | 5 |
| Ca | 12 | 1 | 8 | 8 | 1.00 | 4 | 4 |
| Mg | 15 | 3 | 12 | 1 | 0.19 | 4 | 4 |
| K | 45 | 1 | 24 | 2 | 0.03 | 4 | 4 |
| Mn | –33 | 7 | 1 | 1 | 0.03 | 4 | 4 |
| P | –32 | 2 | –8 | 2 | 0.03 | 4 | 4 |
| N | 4 | 2 | –2 | 7 | 0.47 | 4 | 4 |
| Cорг | 15 | 3 | 12 | 3 | 0.47 | 4 | 4 |
| Лигнин | –80 | 4 | –10 | 3 | 0.03 | 4 | 4 |
| C:N | 48 | 3 | 99 | 8 | 0.03 | 4 | 4 |
| C:P | 538 | 20 | 1365 | 38 | 0.03 | 4 | 4 |
| N:P | 11 | 0.3 | 14 | 1 | 0.03 | 4 | 4 |
| Лигнин:N | 32 | 0.4 | 45 | 3 | 0.03 | 4 | 4 |
| Брусника | |||||||
| Масса | 20 | 1 | 16 | 0.3 | 0.01 | 5 | 5 |
| Ca | 5 | 0.1 | 10 | 4 | 0.31 | 4 | 4 |
| Mg | 5 | 4 | 16 | 1 | 0.03 | 4 | 4 |
| K | 36 | 4 | 35 | 2 | 0.67 | 4 | 4 |
| Mn | 8 | 5 | –2 | 2 | 0.31 | 4 | 4 |
| P | –4 | 7 | 8 | 1 | 0.31 | 4 | 4 |
| N | 2 | 3 | –3 | 1 | 0.31 | 4 | 4 |
| Cорг | 14 | 6 | 7 | 4 | 0.67 | 4 | 4 |
| Лигнин | –172 | 6 | –193 | 16 | 0.67 | 4 | 4 |
| C:N | 44 | 2 | 61 | 5 | 0.03 | 4 | 4 |
| C:P | 578 | 78 | 693 | 42 | 0.67 | 4 | 4 |
| N:P | 13 | 1 | 11 | 0.4 | 0.67 | 4 | 4 |
| Лигнин:N | 42 | 0.4 | 52 | 3 | 0.03 | 4 | 4 |
| Вороника | |||||||
| Масса | 20 | 3 | 17 | 2 | 0.21 | 5 | 5 |
| Ca | 14 | 6 | 6 | 2 | 0.67 | 4 | 4 |
| Mg | 21 | 0.3 | 29 | 2 | 0.03 | 4 | 4 |
| K | 41 | 4 | 53 | 3 | 0.03 | 4 | 4 |
| Mn | 10 | 5 | 9 | 3 | 0.89 | 4 | 4 |
| P | 9 | 8 | 13 | 4 | 0.89 | 4 | 4 |
| N | 10 | 3 | 5 | 6 | 0.31 | 4 | 4 |
| Cорг | 28 | 4 | 12 | 4 | 0.03 | 4 | 4 |
| Лигнин | 7 | 3 | 6 | 1 | 0.89 | 4 | 4 |
| C:N | 40 | 1 | 62 | 7 | 0.03 | 4 | 4 |
| C:P | 499 | 14 | 626 | 59 | 0.11 | 4 | 4 |
| N:P | 12 | 1 | 10 | 0.4 | 0.03 | 4 | 4 |
| Лигнин:N | 30 | 0.01 | 39 | 5 | 0.03 | 4 | 4 |
| Мхи | |||||||
| Масса | 4 | 1 | 10 | 0.4 | 0.01 | 5 | 5 |
| Ca | –7 | 3 | 8 | 10 | 0.31 | 4 | 4 |
| Mg | 11 | 7 | 32 | 12 | 0.31 | 4 | 4 |
| K | 43 | 4 | 54 | 8 | 0.67 | 4 | 4 |
| Mn | 1 | 9 | 35 | 6 | 0.11 | 4 | 4 |
| P | 5 | 4 | 28 | 4 | 0.03 | 4 | 4 |
| N | –11 | 5 | 27 | 7 | 0.03 | 4 | 4 |
| Cорг | –3 | 7 | 12 | 4 | 0.11 | 4 | 4 |
| Лигнин | 2 | 3 | 4 | 6 | 0.89 | 4 | 4 |
| C:N | 69 | 4 | 77 | 0.3 | 0.19 | 4 | 4 |
| C:P | 623 | 35 | 684 | 42 | 0.47 | 4 | 4 |
| N:P | 9 | 0.2 | 9 | 1 | 0.47 | 4 | 4 |
| Лигнин:N | 26 | 1 | 34 | 3 | 0.03 | 4 | 4 |
| 2 год | |||||||
| Хвоя* | |||||||
| Масса | 37 | 1 | 29 | 0.5 | 0.01 | 5 | 5 |
| Ca | 15 | 5 | 4 | 1 | 0.06 | 4 | 4 |
| Mg | 20 | 5 | 29 | 3 | 0.19 | 4 | 4 |
| K | 80 | 1 | 50 | 1 | 0.03 | 4 | 4 |
| Mn | –11 | 14 | 16 | 3 | 0.31 | 4 | 4 |
| P | 10 | 14 | –1 | 2 | 0.89 | 4 | 4 |
| N | 11 | 2 | 19 | 2 | 0.06 | 4 | 4 |
| Cорг | 28 | 4 | 27 | 3 | 0.89 | 4 | 4 |
| Лигнин | –35 | 9 | 5 | 1 | 0.03 | 4 | 4 |
| C:N | 44 | 3 | 103 | 7 | 0.03 | 4 | 4 |
| C:P | 711 | 91 | 1215 | 39 | 0.03 | 4 | 4 |
| N:P | 17 | 3 | 12 | 0.4 | 0.31 | 4 | 4 |
| Лигнин:N | 26 | 2 | 49 | 2 | 0.03 | 4 | 4 |
| Брусника | |||||||
| Масса | 33 | 3 | 26 | 1 | 0.21 | 5 | 5 |
| Ca | 6 | 0.2 | 9 | 1 | 0.03 | 4 | 4 |
| Mg | 9 | 3 | 30 | 1 | 0.03 | 4 | 4 |
| K | 62 | 1 | 74 | 1 | 0.03 | 4 | 4 |
| Mn | 5 | 2 | 2 | 1 | 0.31 | 4 | 4 |
| P | 4 | 1 | 27 | 2 | 0.03 | 4 | 4 |
| N | 4 | 1 | 6 | 3 | 0.67 | 4 | 4 |
| Cорг | 28 | 6 | 19 | 3 | 0.67 | 4 | 4 |
| Лигнин | –131 | 27 | –185 | 17 | 0.31 | 4 | 4 |
| C:N | 38 | 3 | 59 | 3 | 0.03 | 4 | 4 |
| C:P | 510 | 47 | 772 | 31 | 0.03 | 4 | 4 |
| N:P | 13 | 0.2 | 13 | 1 | 0.89 | 4 | 4 |
| Лигнин:N | 36 | 4 | 55 | 5 | 0.03 | 4 | 4 |
| Вороника | |||||||
| Масса | 34 | 3 | 25 | 2 | 0.06 | 5 | 5 |
| Ca | 26 | 6 | 16 | 4 | 0.31 | 4 | 4 |
| Mg | 42 | 3 | 40 | 3 | 0.67 | 4 | 4 |
| K | 61 | 3 | 72 | 2 | 0.11 | 4 | 4 |
| Mn | 14 | 1 | 17 | 2 | 0.31 | 4 | 4 |
| P | 20 | 3 | 30 | 3 | 0.03 | 4 | 4 |
| N | 14 | 2 | 0.3 | 2 | 0.03 | 4 | 4 |
| Cорг | 40 | 4 | 28 | 3 | 0.11 | 4 | 4 |
| Лигнин | 22 | 3 | 13 | 0.3 | 0.03 | 4 | 4 |
| C:N | 35 | 2 | 48 | 2 | 0.03 | 4 | 4 |
| C:P | 459 | 18 | 644 | 40 | 0.03 | 4 | 4 |
| N:P | 13 | 0.1 | 13 | 0.3 | 0.89 | 4 | 4 |
| Лигнин:N | 26 | 0.3 | 34 | 2 | 0.03 | 4 | 4 |
| Мхи | |||||||
| Масса | 9 | 2 | 14 | 1 | 0.04 | 5 | 5 |
| Ca | –18 | 2 | 13 | 9 | 0.03 | 4 | 4 |
| Mg | 20 | 8 | 37 | 12 | 0.67 | 4 | 4 |
| K | 62 | 4 | 71 | 3 | 0.06 | 4 | 4 |
| Mn | –19 | 1 | 33 | 11 | 0.03 | 4 | 4 |
| P | 11 | 2 | 37 | 3 | 0.03 | 4 | 4 |
| N | –6 | 1 | 16 | 17 | 0.89 | 4 | 4 |
| Cорг | –2 | 10 | 23 | 7 | 0.06 | 4 | 4 |
| Лигнин | 21 | 7 | 14 | 5 | 0.31 | 4 | 4 |
| C:N | 72 | 8 | 61 | 4 | 0.11 | 4 | 4 |
| C:P | 655 | 83 | 678 | 15 | 0.67 | 4 | 4 |
| N:P | 9 | 0.2 | 11 | 1 | 0.03 | 4 | 4 |
| Лигнин:N | 22 | 2 | 29 | 4 | 0.31 | 4 | 4 |
Примечание: Е-БГЦ — еловый биогеоценоз, С-БГЦ — сосновый биогеоценоз, * — в Е-БГЦ представлены данные по составу хвои ели, в С-БГЦ — хвои сосны; р — вероятность ошибки 1-го рода в U-тесте Манна-Уитни
Как после первого, так и после второго года разложения, хвоя ели по сравнению с хвоей сосны достоверно быстрее теряла массу (20 против 16% и 37 против 29% соответственно), наблюдались более активные потери K и накопление лигнина (p<0.05) (табл. 2, 3). Кроме того, в первый год разложения у хвои ели наблюдалось более активное накопление P. После первого года у хвои ели были выше коэффициент обогащения для Mn, P и лигнина, после второго года — для Mg, N и лигнина. Коэффициент обогащения для K был выше для хвои сосны как после первого, так и после второго года разложения. Стехиометрические соотношения C:N, C:P и лигнин:N после первого и второго года разложения были ýже у опада хвои ели.
Таблица 3. Коэффициент обогащения активных фракций растительного опада в северотаежных сосновых и еловых лесах
| Параметр | Е-БГЦ | С-БГЦ | р | n | |||
| Среднее | Стандартная ошибка | Среднее | Стандартная ошибка | Е-БГЦ | С-БГЦ | ||
| 1 год | |||||||
| Хвоя* | |||||||
| Ca | 1.10 | 0.01 | 1.09 | 0.08 | 1.00 | 4 | 4 |
| Mg | 1.06 | 0.05 | 1.05 | 0.02 | 1.00 | 4 | 4 |
| K | 0.69 | 0.01 | 0.91 | 0.03 | 0.03 | 4 | 4 |
| Mn | 1.67 | 0.10 | 1.18 | 0.01 | 0.03 | 4 | 4 |
| P | 1.65 | 0.03 | 1.28 | 0.03 | 0.03 | 4 | 4 |
| N | 1.20 | 0.02 | 1.22 | 0.08 | 0.89 | 4 | 4 |
| Cорг | 1.06 | 0.04 | 1.04 | 0.04 | 0.67 | 4 | 4 |
| Лигнин | 2.27 | 0.03 | 1.31 | 0.04 | 0.03 | 4 | 4 |
| Брусника | |||||||
| Ca | 1.19 | 0.02 | 1.07 | 0.04 | 0.11 | 4 | 4 |
| Mg | 1.18 | 0.07 | 1.00 | 0.005 | 0.03 | 4 | 4 |
| K | 0.80 | 0.06 | 0.78 | 0.03 | 0.89 | 4 | 4 |
| Mn | 1.16 | 0.08 | 1.21 | 0.02 | 0.89 | 4 | 4 |
| P | 1.30 | 0.10 | 1.10 | 0.01 | 0.11 | 4 | 4 |
| N | 1.22 | 0.02 | 1.23 | 0.02 | 0.31 | 4 | 4 |
| Cорг | 1.07 | 0.06 | 1.11 | 0.05 | 0.67 | 4 | 4 |
| Лигнин | 3.39 | 0.02 | 3.49 | 0.18 | 0.89 | 4 | 4 |
| Вороника | |||||||
| Ca | 1.06 | 0.04 | 1.13 | 0.04 | 0.67 | 4 | 4 |
| Mg | 0.99 | 0.03 | 0.86 | 0.03 | 0.03 | 4 | 4 |
| K | 0.73 | 0.03 | 0.57 | 0.04 | 0.03 | 4 | 4 |
| Mn | 1.11 | 0.03 | 1.09 | 0.05 | 0.67 | 4 | 4 |
| P | 1.12 | 0.06 | 1.05 | 0.06 | 0.31 | 4 | 4 |
| N | 1.11 | 0.00 | 1.15 | 0.08 | 0.31 | 4 | 4 |
| Cорг | 0.89 | 0.02 | 1.05 | 0.04 | 0.03 | 4 | 4 |
| Лигнин | 1.15 | 0.00 | 1.13 | 0.03 | 0.31 | 4 | 4 |
| Мхи | |||||||
| Ca | 1.12 | 0.03 | 1.02 | 0.10 | 1.00 | 4 | 4 |
| Mg | 0.94 | 0.08 | 0.75 | 0.13 | 0.31 | 4 | 4 |
| K | 0.60 | 0.05 | 0.51 | 0.09 | 0.67 | 4 | 4 |
| Mn | 1.04 | 0.10 | 0.72 | 0.07 | 0.11 | 4 | 4 |
| P | 0.99 | 0.03 | 0.80 | 0.04 | 0.03 | 4 | 4 |
| N | 1.17 | 0.05 | 0.81 | 0.07 | 0.03 | 4 | 4 |
| Cорг | 1.07 | 0.06 | 0.98 | 0.05 | 0.67 | 4 | 4 |
| Лигнин | 1.03 | 0.04 | 1.06 | 0.06 | 0.89 | 4 | 4 |
| 2 год | |||||||
| Хвоя* | |||||||
| Ca | 1.35 | 0.07 | 1.36 | 0.02 | 0.89 | 4 | 4 |
| Mg | 1.28 | 0.07 | 1.00 | 0.03 | 0.03 | 4 | 4 |
| K | 0.32 | 0.01 | 0.70 | 0.02 | 0.03 | 4 | 4 |
| Mn | 1.78 | 0.23 | 1.18 | 0.03 | 0.11 | 4 | 4 |
| P | 1.43 | 0.23 | 1.42 | 0.03 | 0.89 | 4 | 4 |
| N | 1.41 | 0.02 | 1.14 | 0.03 | 0.03 | 4 | 4 |
| Cорг | 1.15 | 0.06 | 1.03 | 0.03 | 0.31 | 4 | 4 |
| Лигнин | 2.16 | 0.14 | 1.34 | 0.02 | 0.03 | 4 | 4 |
| Брусника | |||||||
| Ca | 1.38 | 0.06 | 1.23 | 0.02 | 0.11 | 4 | 4 |
| Mg | 1.33 | 0.02 | 0.95 | 0.02 | 0.03 | 4 | 4 |
| K | 0.55 | 0.01 | 0.35 | 0.02 | 0.03 | 4 | 4 |
| Mn | 1.40 | 0.09 | 1.33 | 0.03 | 0.89 | 4 | 4 |
| P | 1.41 | 0.07 | 0.99 | 0.03 | 0.03 | 4 | 4 |
| N | 1.40 | 0.05 | 1.28 | 0.06 | 0.31 | 4 | 4 |
| Cорг | 1.04 | 0.04 | 1.11 | 0.04 | 0.31 | 4 | 4 |
| Лигнин | 3.34 | 0.24 | 3.86 | 0.20 | 0.31 | 4 | 4 |
| Вороника | |||||||
| Ca | 1.10 | 0.04 | 1.12 | 0.03 | 0.67 | 4 | 4 |
| Mg | 0.87 | 0.01 | 0.80 | 0.03 | 0.11 | 4 | 4 |
| K | 0.59 | 0.02 | 0.38 | 0.02 | 0.03 | 4 | 4 |
| Mn | 1.30 | 0.03 | 1.11 | 0.04 | 0.03 | 4 | 4 |
| P | 1.21 | 0.01 | 0.93 | 0.06 | 0.03 | 4 | 4 |
| N | 1.30 | 0.02 | 1.34 | 0.05 | 0.67 | 4 | 4 |
| Cорг | 0.89 | 0.03 | 0.97 | 0.06 | 0.67 | 4 | 4 |
| Лигнин | 1.17 | 0.00 | 1.17 | 0.03 | 0.89 | 4 | 4 |
| Мхи | |||||||
| Ca | 1.33 | 0.02 | 1.01 | 0.10 | 0.03 | 4 | 4 |
| Mg | 0.90 | 0.09 | 0.74 | 0.14 | 0.89 | 4 | 4 |
| K | 0.42 | 0.04 | 0.34 | 0.04 | 0.31 | 4 | 4 |
| Mn | 1.33 | 0.01 | 0.78 | 0.12 | 0.03 | 4 | 4 |
| P | 1.00 | 0.03 | 0.73 | 0.03 | 0.03 | 4 | 4 |
| N | 1.19 | 0.01 | 0.97 | 0.19 | 0.89 | 4 | 4 |
| Cорг | 1.14 | 0.11 | 0.90 | 0.08 | 0.06 | 4 | 4 |
| Лигнин | 0.88 | 0.08 | 1.00 | 0.06 | 0.31 | 4 | 4 |
Примечание: Е-БГЦ — еловый биогеоценоз, С-БГЦ — сосновый биогеоценоз, * — в Е-БГЦ представлены данные по составу хвои ели, в С-БГЦ — хвои сосны; р — вероятность ошибки 1-го рода в U-тесте Манна-Уитни
Опад листьев брусники в первый год достоверно быстрее разлагался в ельнике, на второй год различия сгладились, но стали более выраженными различия в изменении химического состава опада: коэффициенты обогащения для Mg, K и P были выше в ельнике, и, соответственно, в сосновом лесу листья брусники потеряли больше Ca, Mg, K и P (p<0.05). Соотношения C:N и лигнин:N после первого и второго года были меньше в ельнике. Листья вороники за весь период исследования разлагались в ельниках и сосняках с сопоставимыми скоростями (табл. 2). Но в первый год коэффициенты обогащения Mg и K были выше в ельнике, а потери Mg и K — в сосняке, и, напротив, коэффициент обогащения для С был выше в сосняке, а потери — в ельнике. На второй год коэффициенты обогащения для K, Mn и P были выше в ельнике, при этом опад листьев вороники при разложении терял больше N и лигнина в ельнике, P — в сосняке (p<0.05). В опаде листьев вороники соотношения C:N и лигнин:N на протяжении всего периода исследования были ýже в ельнике.
Выявленные для кустарничков различия могут свидетельствовать о более активных процессах трансформации химического состава опада в ельниках по сравнению с сосновыми лесами, что можно объяснить исходными концентрациями элементов. Так, более высокое исходное содержание Mn в хвое ели и тканях растений напочвенного покрова и накопление его в процессе разложения может ускорять процессы разложения за счет увеличения содержания фермента Mn-пероксидазы, отвечающего за разложение лигнина (Орлова и др., 2011). Повышенное содержание углерода в растительных тканях может свидетельствовать о высоком содержании органических веществ, например лигнина, устойчивых к разложению. По результатам более ранней работы смешанный опад вечнозеленых растений еловых лесов (хвоя ели, листья брусники и вороники) в течение двух лет разлагался быстрее, чем сосновых (хвоя сосны, листья брусники и вороники), что связывали также с различиями качества растительных остатков — более высоким содержанием элементов питания и более узкими соотношениями C:N и лигнин:N в опаде и в почвах еловых лесов (Лукина и др., 2008; Иванова и др., 2019).
Различия в активности процессов разложения в еловых и сосновых лесах могут быть обусловлены не только исходным качеством растительного материала. Основными организмами-деструкторами в бореальных лесах являются сапротрофные грибы, наиболее эффективно разлагающие опад (Hobbie et al., 1999; Bödeker et al., 2016). Согласно литературным данным, общая биомасса микроорганизмов, в т. ч. грибов, выше в еловых биогеоценозах в сравнении с сосновыми (Никонов и др., 2001; Полянская и др., 2001), длина грибного мицелия в органогенных горизонтах почв также больше в еловых лесах (Евдокимова, Мозгова, 2001). Это может быть связано с относительно низкой влажностью почв, формируемой в сосновых лесах (Никонов и др., 2004).
Важным фактором, который влияет на скорость разложения, является температура почв, регулирующая деятельность организмов-деструкторов — низкие температуры ограничивают процессы разложения (Воробьева, Наумова, 2009; Rief et al., 2012). Однако по данным измерений, проведенных в 2015–2021 гг. с помощью температурных логгеров, заложенных под органогенными горизонтами почв, среднемесячные температуры в еловых лесах были ниже, чем в сосновых — разница достигала 2 °С (рис. 1). Это позволяет прийти к заключению, что качество опада может оказывать более существенное влияние на скорость разложения, чем температура почвы.
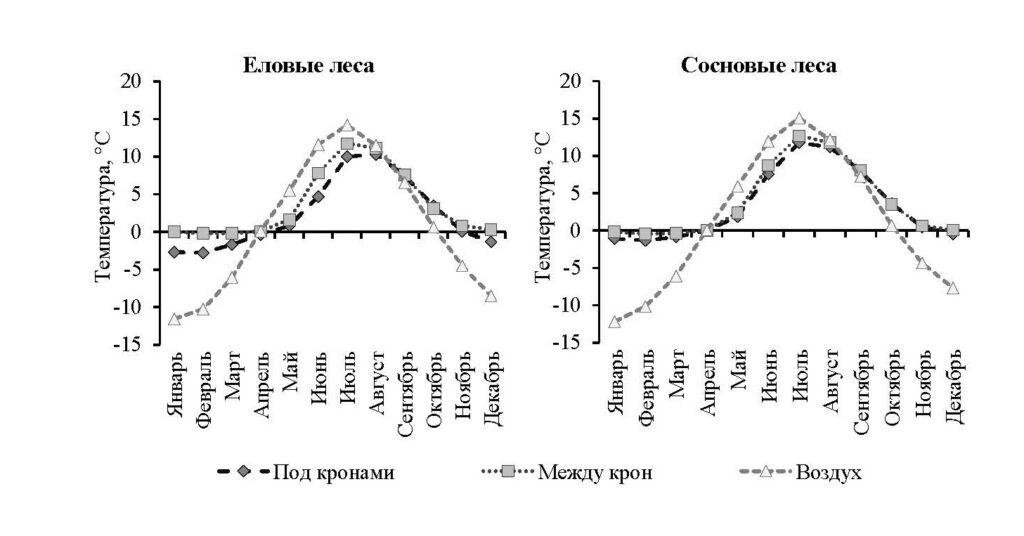
Рисунок 1. Температура почвы и воздуха в еловых и сосновых лесах, измеренные в период 2015–2021 гг. с помощью температурных логгеров под кронами деревьев, в межкроновых пространствах и на деревьях
Отмирающие части зеленых мхов, несмотря на их более высокое качество в ельниках, в отличие от всех остальных видов за весь период разлагались активнее в сосняках (p<0.05) (табл. 2). В первый год коэффициенты обогащения P и N были выше в ельниках, а потери — в сосняках. При этом в ельниках наблюдалось активное накопление N остатками мхов. На второй год зеленые мхи, так же как и в первый, теряли P активнее в сосновом лесу. Возможной причиной может быть большее количество осадков в сосновых лесах — 32.6 мм против 26.5 в ельниках (Ершов, 2021), способствующее механической деструкции растительных остатков мхов и вымыванию подвижных элементов. Кроме того, в сосняке наблюдались потери Ca и Mn, а в ельнике — их аккумуляция; коэффициенты обогащения для Ca, Mn и P были достоверно выше в ельнике. После первого года в ельниках было достоверно ниже соотношение лигнин:N, после второго года — соотношение N:P (табл. 2).
Таким образом, сочетание таких факторов, как качество опада, активность микроорганизмов, температурный режим и количество осадков, связанных с формацией леса, может определять повышенную скорость разложения и интенсивность изменений химического состава растительного материала в еловых лесах по сравнению с сосновыми.
ЗАКЛЮЧЕНИЕ
Результаты исследования показывают, что формация леса оказывает значительное влияние на скорость разложения: в еловых лесах скорость разложения хвои ели выше, чем скорость разложения хвои сосны в сосновых лесах. Существенное влияние на скорость разложения растительных остатков в различных формациях леса оказывало качество растительного материала и, соответственно, активность микроорганизмов. Хвоя ели характеризуется более высоким качеством: содержание Сa, Mg, K, P, Mn выше в стареющей хвое ели по сравнению с содержанием этих элементов в хвое сосны последних лет жизни; отношение C:N, C:P ýже в хвое ели. Это объясняет более активные процессы разложения опада ели в еловых лесах: 37% потерь массы после двух лет разложения в ельнике против 29% — в сосняке. Стареющие, готовые перейти в опад, отмирающие органы кустарничков и мхов проявили неоднозначные различия в содержании элементов питания в зависимости от формации леса: качество живых стареющих листьев брусники и живых побегов мхов было выше в еловых лесах. Тем не менее, в тканях всех видов растений в еловых лесах оказалось больше Mn, тогда как в сосновых лесах — C. Это может быть связано как с условиями местообитания, которые формируются под действием деревьев-эдификаторов (плодородие почв, влажность, температурный режим), так и непосредственным влиянием древесных растений: в сосновых лесах за счет ажурности крон создаются более благоприятные условия освещенности для активного фотосинтеза и накопления углерода растениями напочвенного покрова; повышенное содержание Mn в растениях напочвенного покрова ельников связано с поступлением его в почву с опадом хвои ели, также богатым марганцем. Соответственно этому и различия в скорости разложения опада кустарничков и мхов между формациями лесов проявляются не четко. Скорость разложения листьев брусники была выше в еловых лесах после года разложения, что может быть связано с более высоким качеством растительного материала в еловом лесу: узкими соотношениями элементов (C:N, C:P, N:P) и высоким содержанием элементов питания (Mg, Mn, P). В то же время отмершие части зеленых мхов разлагались активнее в сосновых лесах, несмотря на более высокое их качество в ельниках, что может объясняться большим количеством осадков в сосняках. Таким образом, различия в скорости разложения растительных остатков хвои древесных растений, листьев кустарничков и побегов мхов определяется сочетанием таких факторов, как качество растительного материала, температурный режим и количество осадков, связанных с формацией леса.
БЛАГОДАРНОСТИ
Авторы благодарят Артемкину Наталью Александровну, старшего научного сотрудника лаборатории наземных экосистем Института проблем промышленной экологии Севера ФИЦ КНЦ РАН, за проведение анализов на содержание лигнина в образцах опада для экспериментов по разложению на площадках мониторинговой сети ИППЭС КНЦ РАН.
ФИНАНСИРОВАНИЕ
Исследование выполнено в рамках работы молодежной лаборатории ЦЭПЛ РАН «Климаторегулирующие функции и биоразнообразие лесов» (регистрационный номер 122111500023-6).
СПИСОК ЛИТЕРАТУРЫ
Аткина Л. И. Географо-лесотипологические закономерности структуры и запаса напочвенного покрова таежных лесов: автореф. дисс. докт. с-х. наук (спец. 06.03.03). Екатеринбург, 2000. 37 с.
Атлас Мурманской области. М.: Изд-во Главного Управления Геодезии и Картографии при Совете Министров СССР, 1971. 44 с.
Белов Н. П., Барановская А. В. Почвы Мурманской области. Л.: Наука, 1969. 147 с.
Бобкова К. С. Роль лесной подстилки в функционировании хвойных экосистем Европейского Севера // Вестник Института биологии Коми НЦ УрО РАН. 2000. № 9 (35). URL: https://kurl.ru/oMrnF (дата обращения 15.11.2023).
Воробьева И. Г., Наумова А. Н. Интенсивность процесса деструкции растительного опада в почвах сухих местообитаний // Продуктивность и устойчивость лесных почв: Матер. III междунар. конф., г. Петрозаводск, 7‒11 сентября 2009 г. Петрозаводск: Изд-во Карельского НЦ РАН, 2009. C. 192–195.
Воробьева Л. А. Химический анализ почв: Учебник. М.: Изд-во Московского государственного университета, 1998. 272 с.
Евдокимова Г. А., Мозгова Н. П. Микроорганизмы тундровых и лесных подзолов Кольского Севера. Апатиты: КНЦ РАН, 2001. 184 с.
Ершов В. В. Фитогенное варьирование состава атмосферных выпадений и почвенных вод северотаежных лесов в условиях аэротехногенного загрязнения: Дисс. … канд. биол. наук (спец. 1.5.15). Апатиты, 2021. 188 с.
Зубкова Е. В., Фролов П. В., Быховец С. С., Надпорожская М. А., Фролова Г. Г. Мозаичность ценопопуляций черники и брусники, и динамика органического вещества почвы в сосняках Южного Подмосковья // Лесоведение. 2022. № 1. С. 34–46.
Иванова Е. А., Лукина Н. В., Данилова М. А., Артемкина Н. А., Смирнов В. Э., Ершов В. В., Исаева Л. Г. Влияние аэротехногенного загрязнения на скорость разложения растительных остатков в сосновых лесах на северном пределе распространения // Лесоведение. 2019. № 6. С. 533–546.
Исаева Л. Г., Сухарева Т. А. Элементный состав дикорастущих кустарничков в зоне воздействия комбината «Североникель»: данные многолетнего мониторинга // Цветные металлы. 2013. № 10. С. 87–92.
Кищенко И. Т. Лесоведение и лесная экология. Учебное пособие для бакалавриата и магистратуры. М.: Изд-во Юрайт, 2019. 393 с.
Колмогорова Е. Ю., Уфимцев В. И. Некоторые особенности химического состава опада сосны обыкновенной, произрастающей в условиях породного отвала // Успехи современного естествознания. 2018. № 11. Ч. 2. С. 267–272.
Кузнецов М. А. Влияние условий разложения и состава опада на характеристики и запас подстилки в среднетаежном чернично-сфагновом ельнике // Лесоведение. 2010. № 6. С. 54–60.
Кузнецов М. А., Осипов А. Ф. Растительный опад как компонент биологического круговорота углерода в заболоченных хвойных сообществах средней тайги // Вестник Института биологии Коми НЦ УрО РАН. 2011. № 9. С. 10–12.
Ларионова А. А., Квиткина А. К., Быховец С. С., Лопес де Гереню В. О., Колягин Ю. Г., Каганов В. В. Влияние азота на минерализацию и гумификацию лесных опадов в модельном эксперименте // Лесоведение. 2017. № 2. С. 128–139.
Лукина Н. В., Горбачева Т. Т., Никонов В. В., Лукина М. А. Пространственная изменчивость кислотности AL-Fe-гумусовых подзолов // Почвоведение. 2002. № 2. С. 163–176.
Лукина Н. В., Никонов В. В. Биогеохимические циклы в лесах Севера в условиях аэротехногенного загрязнения: в 2-х ч. Апатиты: Изд-во КНЦ РАН, 1996. Ч. 1. 213 с.; Ч. 2. 192 с.
Лукина Н. В., Никонов В. В. Питательный режим лесов северной тайги: природные и техногенные аспекты. Апатиты: Изд-во КНЦ РАН, 1998. 316 с.
Лукина Н. В., Никонов В. В., Исаева Л. Г. Кислотность и питательный режим почв еловых лесов // Коренные еловые леса: биоразнообразие, структура, функции. СПб.: Наука, 2006. 298 с.
Лукина Н. В., Орлова М. А., Исаева Л. Г. Плодородие лесных почв как основа взаимосвязи почва — растительность // Лесоведение. 2010. № 5. С. 45–56.
Лукина Н. В., Полянская Л. М., Орлова М. А. Питательный режим почв северотаежных лесов. М.: Наука, 2008. 342 с.
Молчанов А. Г. Сравнение экофизиологических показателей сосны и ели в Серебряноборском опытном лесничестве // Лесохозяйственная информация. 2020. № 1. С. 115–124.
Никонов В. В., Лукина Н. В., Безель В. С., Бельский Е. А., Беспалова А. Ю., …, & Яценко-Хмелевская М. А. Рассеянные элементы в бореальных лесах. М.: Наука, 2004. 616 с.
Никонов В. В., Лукина Н. В., Полянская Л. М., Паникова А. Н. Особенности распространения микроорганизмов в Al-Fe-гумусовых подзолах северотаежных еловых лесов: природные и техногенные аспекты // Микробиология. 2001. Т. 70. № 3. С. 319‒328.
Орлова М. А., Лукина Н. В., Камаев И. О., Смирнов В. Э., Кравченко Т. В. Мозаичность лесных биогеоценозов и продуктивность почв // Лесоведение. 2011. № 6. С. 39–48.
Переверзев В. Н. Лесные почвы Кольского полуострова. М.: Наука, 2004. 232 с.
Полянская Л. М., Никонов В. В., Лукина Н. В., Паникова А. Н., Звягинцев В. Г. Микроорганизмы Al-Fe-гумусовых подзолов сосняков лишайниковых в условиях аэротехногенного загрязнения // Почвоведение. 2001. № 2. С. 215‒226.
Помогайбин Е. А., Помогайбин А. В. Влияние деревьев рода Juglans L. на целлюлозоразрушающую активность почвы в условиях дендрария ботанического сада Самарского университета // Самарский научный вестник. 2018. Т. 7. № 1 (22). С. 105–109.
Рахлеева А. А., Семенова Т. А., Стриганова Б. Р., Терехова В. А. Динамика зоомикробных комплексов при разложении растительного опада в ельниках южной тайги // Почвоведение. 2011. № 1. С. 44–55.
Суворова Г. Г. Фотосинтетическая активность хвойных деревьев в условиях юга Средней Сибири: автореф. дисс. … докт. биол. наук (спец. 03.00.12). Иркутск, 2006. 40 с.
Сухарева Т. А. Элементный состав зеленых мхов фоновых и техногенно нарушенных территорий // Ученые записки Петрозаводского государственного университета. 2018. № 3 (172). С. 89–96.
Сухарева Т. А., Лукина Н. В. Минеральный состав ассимилирующих органов хвойных деревьев после снижения уровня атмосферного загрязнения на Кольском полуострове // Экология. 2014. № 2. С. 97–104.
Тужилкина В. В. Фотосинтетическая активность сосны и ели в условиях средней подзоны тайги Коми АССР: Дисс. … канд. биол. наук (спец. 03.00.12). Сыктывкар, 1984. 148 с.
Федорец Н. Г., Бахмет О. Н. Особенности формирования почв и почвенного покрова Карело-Кольского региона // Труды Карельского научного центра РАН. 2016. № 12. С. 39‒51.
Цандекова О. Л. Динамика накопления золы в опаде Acer negundo L. в условиях нарушенных пойменных фитоценозов // Бюллетень науки и практики. 2018. Т. 4. № 12. С. 148‒152.
Цветков В. Ф. Лесной биогеоценоз (2-е издание, исправленное и дополненное). Архангельск, 2004. 267 с.
Чертов О. Г., Меньшикова Г. П. Изменение лесных почв под действием кислых осадков // Известия АН СССР. Серия биология. 1983. № 6. С.110–115.
Aponte C., García L. V., Marañón T. Tree species effects on nutrient cycling and soil biota: A feedback mechanism favouring species coexistence // Forest Ecology and Management. 2013. Vol. 309. P. 36–46.
Berg B. Litter decomposition and organic matter turnover in northern forest soils // Forest Ecology and Management. 2000. Vol. 133. P. 13–22.
Berg B., McClaugherty C. Plant litter — decomposition, humus formation, carbon sequestration, 2nd ed. Germany: Springer-Verlag Berlin Heidelberg, 2008. 340 p.
Bödeker I. T. M., Lindahl B. D., Olson Å., Clemmensen K. E. Mycorrhizal and saprotrophic fungal guilds compete for the same organic substrates but affect decomposition differently // Functional Ecology. British Ecological Society. 2016. Vol. 30. No 12. P. 1967–1978.
Bradford M. A., Berg B., Maynard D. S., Wieder W. R., Wood S. A. Understanding the dominant controls on litter decomposition // Journal of Ecology. 2016. Vol. 104. No 1. P. 229–238.
Chavez-Vergara B., Merino A., Vázquez-Marrufo G., García-Oliva F. Organic matter dynamics and microbial activity during decomposition of forest floor under two native neotropical oak species in a temperate deciduous forest in Mexico // Geoderma. 2014. Vol. 235–236. P. 133–145.
Cornelissen J. H. C., van Bodegom P. M., Aerts R., Callaghan T. V., van Logtestijn R. S. P., …, & Team M. O. L. Global negative vegetation feedback to climate warming responses of leaf decomposition rates in cold biomes // Ecology Letters. 2007. Vol. 10. P. 619–627.
De Marco A., Vittozzi P., Rutigliano F. A., Virzo de Santo A. Nutrient dynamics during decomposition of four different pine litters // V. Leone, R. Lovreglio (Eds.). Proceedings of the International Workshop MEDPINE 3: Conservation, Regeneration and Restoration of Mediterranean Pines and Their Ecosystems. Bari: CIHEAM, 2007. P. 73–77.
Fang X., Zhao L., Zhou G., Huang W., Liu J. Increased litter input increases litter decomposition and soil respiration but has minor effects on soil organic carbon in subtropical forests // Plant and Soil. 2015. Vol. 392. P. 139–153.
Helmisaari H.-S. Temporal variation in nutrient concentrations of Pinus sylvestris needles // Canadian Journal of Forest Research. 1990. No 5. P. 177–193.
Hilli S. Significance of litter production of forest stands and ground vegetation in the formation of organic matter and storage of carbon in boreal coniferous forests // Forest Condition Monitoring in Finland — National report / P. Merilä, S. Jortikka (Eds.) The Finnish Forest Research Institute. 2013. P. 77–83.
Hobbie E. A., Macko S. A., Shugart H. H. Insights into nitrogen and carbon dynamics of ectomycorrhizal and saprotrophic fungi from isotopic evidence // Oecologia. 1999. Vol. 118. No 3. P. 353–360.
Högberg P., Näsholm T., Franklin O., Högberg M. N. Tamm Review: On the nature of the nitrogen limitation to plant growth in Fennoscandian boreal forests // Forest Ecology and Management. 2017. Vol. 403. P. 161–185.
Husson F., Le S., Pages J. Exploratory Multivariate Analysis by Example Using R. 2nd Ed. London: Chapman & Hall/CRC Press. 2017. 248 p.
Leonard L. T., Mikkelson K., Hao Z., Brodie E. L., Williams K. H., Sharp J. O. A comparison of lodgepole and spruce needle chemistry impacts on terrestrial biogeochemical processes during isolated decomposition // PeerJ. 2020. Vol. 8. Article: e9538.
Ligrone R., Carafa A., Duckett J. G., Renzaglia K. S., Ruel K. Immunocytochemical detection of lignin-related epitopes in cell walls in bryophytes and the charalean alga Nitella // Plant Systematics and Evolution. 2008. Vol. 270. P. 257–272.
Lukina N. V., Orlova M. A., Steinnes E., Artemkina N. A., Gorbacheva T. T., Smirnov V. E., Belova E. A. Mass-loss rates from decomposition of plant residues in spruce forests near the northern tree line subject to strong air pollution // Environmental Science and Pollution Research. 2017. Vol. 24. No 24. P. 19874–19887.
Pausas J. G. Litter fall and litter decomposition in Pinus sylvestris forests of the eastern Pyrenees // Journal of Vegetation Science. 1997. Vol. 8. P. 643–650.
Portillo-Estrada M., Pihlatie M., Korhonen J. F. J., Levula J., Frumau A. K. F., Ibrom A., Lembrechts J. J., Morillas L., Horváth L., Jones S. K., Niinemets Ü. Climatic controls on leaf litter decomposition across European forests and grasslands revealed by reciprocal litter transplantation experiments // Biogeosciences. 2016. Vol. 13. P. 1621–1633.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria, 2017. URL: http://www.R-project.org (дата обращения 15.11.2023)
Rahman M. M., Tsukamoto J., Rahman M. M., Yoneyama A., Mostafa K. M. Lignin and its effects on litter decomposition in forests ecosystems // Chemistry and Ecology. 2013. Vol. 29. No 6. P. 540–553.
Rautio P., Huttunen S., Lamppu J. Seasonal foliar chemistry of northern Scots pine under sulphur and heavy metal pollution // Chemosphere. 1998. Vol. 37. No 2. P. 271–287.
Rief A., Knapp B. A., Seeber J. Palatability of selected alpine plant litters for the decomposer Lumbricus rubellus (Lumbricidae) // PLoS One. 2012. Vol. 7. Article: e45345.
Rowland A. P., Roberts J. D. Lignin and cellulose fractionation in decomposition studies using acid-detergent fibre methods // Communications in Soil Science and Plant Analysis. 1994. Vol. 25. No 3–4. P. 269–277.
Tu L-h., Hu H-l., Chen G., Peng Y., Xiao Y-l., Hu T-x., Zhang J., Li X-w., Liu L., Tang Y. Nitrogen addition significantly affects forest litter decomposition under high levels of ambient nitrogen deposition // PLoS ONE. 2014. Vol. 9. No 2. P. 1–9.
Wardle D. A., Nilsson M.-C., Zackrisson O., Gallet C. Determinants of litter mixing effects in a Swedish boreal forest // Soil Biology and Biochemistry. 2003. Vol. 35. No 6. P. 827–835.
Yang Q., Blanco N. E., Hermida-Carrera C., Lehotai N., Hurry V., Strand Å. Two dominant boreal conifers use contrasting mechanisms to reactivate photosynthesis in the spring // Nature Communications. 2020. Vol. 11. Article 128.
Zhang D., Hui D., Luo Y., Zhou G. Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors // Journal of Plant Ecology. 2008. Vol. 1. No 2. P. 85–93.
Рецензент: к. б. н., с. н. с. Ахметова Г. В.





