- DOI: 10.31509/2658-607x-202583-174
- УДК 631.48
Влияние валежных стволов Picea аbies (L.) H. Karst. на строение почвенного профиля
![]()
© 2025 Е. В. Фомина*, А. В. Кикеева, И. В. Ромашкин, А. Ю. Нуколова,
А. М. Крышень
Институт леса Карельского научного центра РАН, ул. Пушкинская, 11, Петрозаводск, Республика Карелия, 185910 Россия
*E-mail: evfomina@krc.karelia.ru
Поступила в редакцию: 30.06.2025
После рецензирования: 25.08.2025
Принята к печати: 09.09.2025
Актуальность и цели. Валежная древесина является неотъемлемым компонентом естественных лесных экосистем, обеспечивающим их устойчивость. В отечественном почвоведении роль крупных древесных остатков как компонента почвенного профиля практически не учитывается, несмотря на то, что погребенная валежная древесина широко представлена в бореальных лесах. Цель исследования – охарактеризовать изменение морфологических свойств (цвет и строение профиля) подзола иллювиально-железистого (Ferric Albic Podzol Arenic) и серогумусовой (дерновой) метаморфизованной почвы (Umbrisol Loamic) под влиянием валежных стволов Picea abies 3-5 классов разложения в локальных условиях среднетаежного ельника. Материалы и методы. Исследование проведено летом 2023 и 2024 гг. в Республике Карелия на территории Государственного природного заповедника «Кивач» на двух пробных площадях. Влияние ствола прослежено от 3 к 5 классу разложения. Почвенные разрезы выкопаны в каждом направлении от валежного ствола до 1 м. Морфологическое описание разреза проведено непосредственно под стволом, на удалениях 0-25, 25-50 и 50-100 см от ствола согласно общепринятым методикам. В настоящей работе показано влияние валежа на почву в пределах локальной территории, и отсутствует экстраполяция полученных результатов на весь ареал почвенного типа. Результаты. Составлены схемы изменения морфологического строения верхних горизонтов под валежным стволом и на разном расстоянии от него. Под валежными стволами в условиях серогумусовой почвы отсутствует горизонт AY1, интенсивно прокрашенный гумусом. В условиях подзола под валежным стволом диагностируется ферментативно-гумусированный горизонт OFHh. В серогумусовой почве на расстоянии 25 см от ствола наблюдается изменение состава лесной подстилки. В строении подзола иллювиально-железистого под стволом последнего класса разложения и на расстоянии до 25 см от него диагностируется отбеленный горизонт Е. Непосредственно на валеже образуется подстилка, которая по мере разложения ствола постепенно соединяется с почвенной подстилкой. Заключение. Валеж 3-5 классов разложения создает неоднородность в поверхностных горизонтах. Влияние на морфологические свойства проявляется в изменении цвета, а также в появлении или наоборот, отсутствии некоторых почвенных горизонтов. Влияние валежа поздних классов разложения (3, 4, 5а классы для серогумусовой почвы и 3, 4 классы – для подзола) проявляется непосредственно под стволом. Влияние валежного ствола распространяется на окружающее пространство только на последнем классе разложения ствола (5б класс для серогумусовой почвы и 5 класс – для подзола).
Ключевые слова: Заповедник Кивач, Albic Podzol Arenic, Umbrisol Loamic, подзол иллювиально-железистый, серогумусовая (дерновая) метаморфизованная почва, морфология почв, почвенный объект, крупные древесные остатки.
Валежная древесина является центром биогеохимических и физических процессов, которые влияют на химический состав (Sokołowski et al., 2024), строение, морфологические свойства почв (Klinka et al., 1995) и биоразнообразие (Harmon et al., 1986). Крупные древесные остатки (КДО) рассматриваются как неотъемлемый компонент естественных лесных экосистем, обеспечивающий их устойчивость (Стороженко, 2011; Шорохова, 2020). Валеж – один из основных источников почвенного органического вещества, запускает каскад реакций образования, трансформации и миграции водорастворимых органических веществ (Яшин и др., 2018). Эти вещества, в свою очередь, инициируют микробиологические процессы (Shannon et al., 2022), определяя биологическую активность (Nazari et al., 2023) и структуру почвенного покрова (Stutz et al., 2019). Несмотря на растущее число исследований, связанных с изучением КДО (Dhar et al., 2022; Nazari et al., 2023; Shi et al., 2024; Wijas et al., 2025), знания об их влиянии на почву ограничены.
В европейской классификации гумусовых форм валеж заключительной стадии разложения определяется как лигноформ или гиперлигниновый парагумус (Zanella et al., 2011; Jabiol et al., 2013). В другой классификации форм гумуса в зависимости от ключевого влияния грибов и/или бриофитов валеж включается в состав Lignomoder или Lignomor (Klinka et al., 1990; Green et al., 1993). В канадской классификации, где уделяется особое внимание образовавшимся в результате разложения валежа горизонтам, выделяют отдельную группу почв – лигниновых фолисолей (Lignic Folisol). Здесь горизонты лесных подстилок OF и OH состоят преимущественно из средне- или хорошо разложившихся древесных материалов (The Canadian system …, 1998). В отечественном почвоведении до сих пор обсуждается возможность рассмотрения изменения почв под валежной древесиной в качестве элемента почвообразовательного процесса, в ходе которого происходит изменение локального биоценоза и микроклимата (Васенев, 1987, 2003; Васенев, Таргульян, 1995). В современной классификации почв России (Шишов и др., 2004) роль КДО как компонента почвенного профиля практически не учитывается. Между тем, в бореальных лесах под лесной подстилкой широко представлена погребенная валежная древесина (Hagemann et al., 2010; Moroni et al., 2010), которая не учитывается ни при проведении лесотаксационных работ (Швиденко и др., 2009), ни как элемент почвообразования (Герасимова и др., 2019).
Многовековая эволюция почв в условиях таежных ландшафтов проявляется почвенными сукцессиями, которые запускаются периодическими импактными нарушениями (Васенев, 2003). Ветровальные нарушения являются одним из ключевых элементов формирования и развития подзолистых почв, в результате которых активизируются факторы почвообразования (концепция таежного ветровального педогенеза) (Васенев, 1987). Последствия массовых ветровалов прослеживаются в зависимости от условий на протяжении десятилетий, столетий (Ulanova, 2000) и даже тысячелетий (Schaetzl, Follmer, 1990). Структура малонарушенных еловых лесов является результатом выпадений, в том числе непрерывных и нерегулярных, отдельных деревьев, (Edman et al., 2007; Fraver et al., 2008; Caron et al., 2009; Hytteborn, Verwijst, 2011; Kuuluvainen et al., 2014), в результате чего поддерживается постоянная гетерогенность среды. Отдельно выпадающие деревья могут по-разному влиять на содержание питательных веществ в почве (Klinka et al., 1990), а также способствуют изменению химических свойств лесной подстилки (Klinka et al., 1995). В настоящее время все больше внимания уделяется исследованиям динамики химических и физических свойств почв под влиянием разлагающейся древесины (Spears et al., 2003; Радюкина, 2004; Gonzalez-Polo et al., 2013; Ханина и др., 2022; Khan et al., 2022). Влияние валежа способно распространяться на окружающее пространство, проявляясь в изменении физико-химических свойств почв на определённой дистанции. Расстояние, на котором прослеживается влияние валежного ствола, зависит от степени его разложения (Stutz et al., 2019), структуры древостоя (Ma et al., 2014), свойств почвы (Shannon et al., 2022), типа почвообразующей породы (Ulyshen et al., 2016).
Влияние КДО на морфологическое строение почвенного профиля до сих пор изучено фрагментарно (Wilding et al., 1983; Васенев, Просвирина, 1988; Khan et al., 2022), что затрудняет оценку их роли в почвообразовательном процессе в бореальных лесах. Имеющиеся данные о влияния валежа на морфологию почв затрагивают только локальное воздействие — зону непосредственно под валежным стволом (Klinka et al., 1995). В условиях сравнительно низкой скорости биологического круговорота в бореальных лесах биогенно-аккумулятивные процессы в системе «КДО – почва» особенно важны. Цель нашего исследования –охарактеризовать морфологические свойства (цвет и строение профиля) контрастно отличающихся почв (подзола иллювиально-железистого и серогумусовой (дерновой) метаморфизованной почвы) и оценить их изменение под влиянием валежных стволов P. abies 3-5 классов разложения в локальных условиях среднетаежного ельника.
ОБЪЕКТЫ И МЕТОДЫ
Исследование провели в летний период 2023–2024 гг. в Республике Карелия на территории Государственного природного заповедника «Кивач». Важным условием было проведение работ в ельнике на одном участке. Работы выполнены на двух пробных площадях (ПП), характеристики которых представлены в таблице. В ельнике черничном (ПП №1: N 62°28′ E 33°96′) представлен подзол иллювиально-железистый (Ferric Albic Podzol Arenic), в ельнике кислично-черничном (ПП №2: N 62°28′ E 33°97′) – серогумусовая (дерновая) метаморфизованная почва (Umbrisol Loamic). В пределах каждой ПП были подобраны валежные стволы P. abies 3-5 классов разложения, располагающиеся в сходных условиях (рис. 1). Таким образом, влияние валежного ствола прослежено от 3 к 5 классу разложения в однократной повторности для двух типов почв. Закладка большего числа повторностей затруднительна из-за вариабельности условий местообитаний. В настоящей работе показано влияние валежа на почву в пределах локальной территории, что не позволяет экстраполировать полученные результаты на весь ареал распространения рассмотренных типов почв. Поэтому далее в тексте при описании и интерпретации результатов используется термин «почвенный объект» (Козлов, 2003; Самсонова, Мешалкина, 2020).
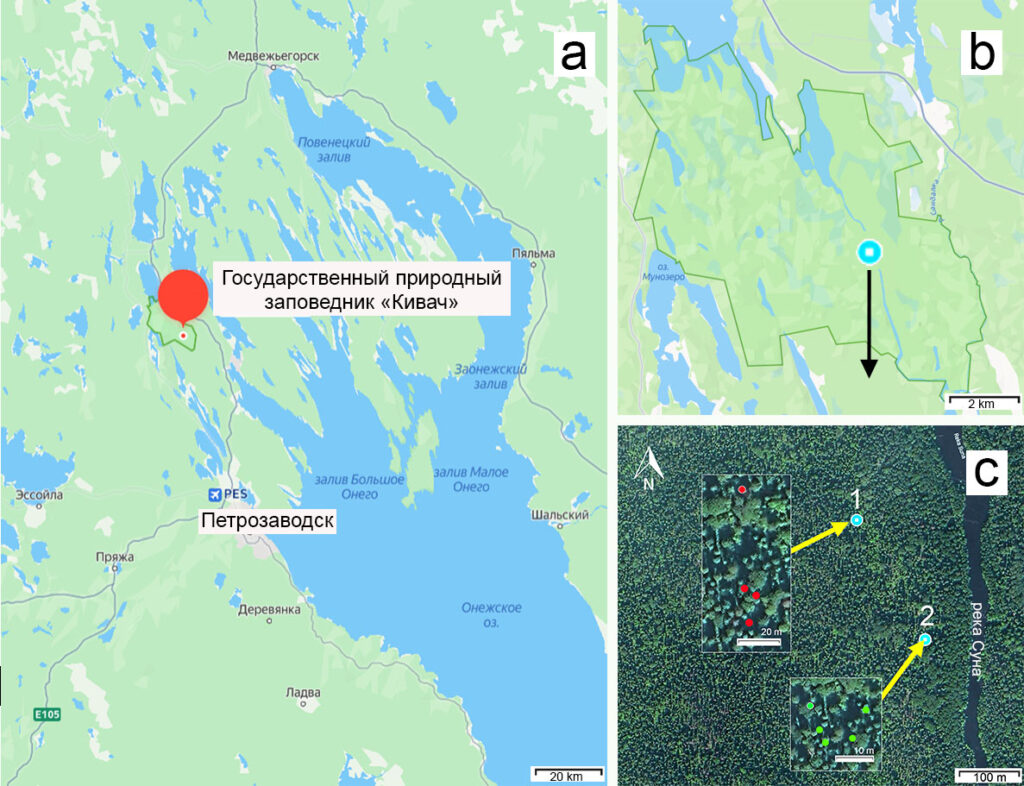
Рисунок 1. Район исследования: a, b – положение заповедника «Кивач» на карте , с – расположение пробных площадей на территории заповедника: 1 – пробная площадь ПП №1; 2 – пробная площадь ПП №2. Красными точками обозначено расположение почвенных объектов на ПП №1, зелеными – обозначено расположение почвенных объектов на ПП №2.
Класс разложения древесины валежных стволов определяли в соответствии с пятистадийной классификацией (Shorohova, Kapitsa, 2015). Для серогумусовой (дерновой) метаморфизованной почвы валежные стволы 5 класса разложения разделяли на два подкласса – 5а и 5б. Стволы 5а класса имели визуально идентифицируемые контуры, 5б класса – практически сливались с поверхностью почвы и имели сложно или практически не различимые контуры. На ПП с подзолом иллювиально-железистым стволы 5 класса разложения хорошо визуализировались. Далее в тексте стволы 3, 4, 5а классов для ПП с серогумусовой почвы и 3, 4 классов для ПП с подзолом обозначали поздними классами разложения, стволы 5б класса для серогумусовой почвы и 5 класса для подзола – последним классом разложения.
Характеристика пробных площадей
| № | Площадь,
га |
Тип леса | Состав и возраст
Древостоя |
Запас,
м3/га |
Полнота | Запас
КДО, м3/га |
Тип почвы |
| 1 | 0.30 | ельник черничный
(Piceetum myrtillosum |
8Е1801Б601Ос70 | 340 | 0.8 | 97 | подзол иллювиально- железистый
O–Ehi–E–BHF–Cg |
| 2 | 0.38 | ельник
кислично-черничный (Piceetum oxalidoso—myrtillosum) |
6Е1603Ос801Б80 | 319 | 0.8 | 162 | серогумусовая метаморфизованная почва
O–AY–Cm–С |
Примечание: КДО – крупные древесные остатки
В связи с тем, что влияние валежного ствола прослеживается на протяжении до 1 м от него (Gonzalez-Polo et al., 2013; Piaszczyk et al., 2019; Błońska, Lasota, 2023), почвенные разрезы закладывали в виде траншеи по обе стороны от валежного ствола на расстояние до 1 м (рис. 2). Почвенные разрезы (траншеи) закладывали в межкроновых пространствах не ближе 1.5 м к стволам живых деревьев в условиях выровненного рельефа и в автоморфных условиях. Траншеи располагали перпендикулярно валежным стволам, глубина разреза – до почвообразующей породы. Морфологическое описание профилей проводили непосредственно под стволом, на удалении 0-25 см, 25-50 и 50-100 см от ствола согласно общепринятым методикам (Острикова, 2008; Шишов и др., 2004). Контрольные почвенные разрезы закладывали на расстоянии не менее 1 м от валежных стволов. Фиксировали морфологические свойства почвенных объектов – мощность, строение, плотность, цвет, новообразования, включения, влажность, степень каменистости, степень насыщенности корнями и гранулометрический состав методом шнура. Мощности почвенных горизонтов фиксировали сантиметровой лентой. Цвет горизонтов определяли по цветовой почвенной шкале Манселла (Munsell, 2000). Фотографии разрезов сводили из совокупности соответствующих указанным расстояниям снимков с использованием программ Adobe Photoshop, Adobe InDesign, а также AutoCAD. Для идентификации типа/подтипа почв использовали классификацию почв России (Шишов и др., 2004) и международную почвенную классификацию – WRB (World Reference …, 2014).

Рисунок 2. Схема закладки почвенного разреза. Цифрами обозначены расстояния от валежного ствола (см).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Описание почвенных разрезов, заложенных вне зоны влияния валежных стволов, показало, что почвы выбранных пробных площадей характеризуются типичным строением и сочетанием генетических горизонтов. В ельнике кислично-черничном (ПП № 2) рассмотрен контрольный разрез серогумусовой (дерновой) метаморфизованной почвы (рис. 3 a). Данный тип почвы сформирован на озерно-ледниковых глинистых и суглинистых отложениях при участии травянистой растительности в относительно богатых флористических условиях (большее количество видов). Разрез заложен в средней части террасы cо слабым уклоном около 3-4º северной экспозиции на левом берегу р. Суна. В напочвенном покрове преобладают ландыш майский (Convallaria majalis L.), майник двулистный (Maianthemum bifolium (L.) F.W. Schmidt), земляника лесная (Fragaria vesca L.), костяника (Rubus saxatilis L.), кислица обыкновенная (Oxalis acetosella L.), единично встречается черника обыкновенная (Vaccinium myrtillus L.). Профиль серогумусовой (дерновой) метаморфизованной почвы включает три генетических горизонта и имеет строение: O–AY–Cm–С. Горизонт лесной подстилки O представлен одним подгоризонтом OL, маломощный (до 1 см), состоит из слаборазложившегося опада. Диагностический серогумусовый горизонт AY отчетливо разделяется на два слоя – AY1 и AY2. AY1 имеет буровато-серый цвет, среднесуглинистый, структура комковатая с элементами зернистости за счет копролитов дождевых червей. AY2 – серый с палевыми пятнами слой, среднесуглинистый, структура комковатая. Следует отметить, что верхний горизонт AY1 сильнее прокрашен гумусом и имеет по шкале Манселла оттенки 10 YR 2/2; 10 YR 3/1 во влажном состоянии и 10 YR 5/1 в сухом состоянии (Munsell, 2000). В то же время AY2 характеризуется более светлыми оттенками: 2.5 Y 3/2; 2.5 Y 5/2 во влажном состоянии и 2.5 Y 7/2 в сухом состоянии, что связано с меньшей прокраской гумусовыми веществами. Горизонт AY1 имеет более зернистую (крупитчатую) структуру, AY2 – комковатую. Кроме того, в нем больше фрагментов нижележащего горизонта, благодаря чему формируется пятнистая (палевые и серые тона) окраска. Горизонт Cm – сизовато-палевый с рыжеватыми пятнами (2.5Y 6/1 во влажном состоянии и 2.5Y 6/3 в сухом состоянии; 2.5Y 4/3 во влажном состоянии и 2.5Y 6/2 в сухом состоянии), уплотненный, тяжелосуглинистый, имеет комковато-глыбистую структуру и постепенно сменяется почвообразующей породой С. Горизонт Cm является переходным к почвообразующей породе и имеет слабые признаки структурного метаморфизма, выражающиеся в наличии признаков оструктуренности, что свидетельствует о слабых процессах педогенного преобразования почвообразующей породы. Горизонт С морфологически разделяется на два подгоризонта: C1 и C2. Горизонт С1 сизовато-палевый со светло-рыжими пятнами (2.5Y 6/1 во влажном состоянии и 2.5Y 6/3 в сухом состоянии; 2.5Y 5/4 во влажном состоянии и 2.5Y 6/3 в сухом состоянии), плотный, тяжелосуглинистый, глыбистой структуры. Горизонт С2 сизовато-палевый (10YR 6/3 во влажном состоянии и 10YR 7/2 в сухом состоянии), плотнее предыдущего.
Разрез подзола иллювиально-железистого супесчаного на двучленных озерно-ледниковых отложениях (песок мелкий, подстилаемый тяжелым суглинком) (рис. 3 b) заложен на ПП №1 в ельнике черничном. Данный тип почвы является типичным для бореальных лесов и формируется во флористически бедных условиях, для которых характерно малое количество видов напочвенного покрова, на песчаных и супесчаных озерно-ледниковых отложениях. Разрез заложен в средней части террасы cо слабым уклоном около 4º северной экспозиции на левом берегу р. Суна. В напочвенном покрове преобладают черника обыкновенная (Vaccinium myrtillus L.), вейник лесной (Calamagrostis arundinacea (L.) Roth), брусника (Vaccinium vitis-idaea L.). На поверхности почвы и в верхней части профиля (0-25 см) часто встречаются крупные камни-валуны. Профиль подзола иллювиально-железистого включает пять генетических горизонтов и имеет строение O–Ehi–E–BHF–Cg. Подстилочно-торфяный горизонт O мощностью до 8 см дифференцирован на два слоя – ОL и OF. Горизонт OL (подгоризонт свежего опада) характеризуется наличием опада текущих и прошлых лет, который дифференцируется в основном на листья, хвою, ветки. Горизонт OF (подгоризонт ферментации) состоит из полуразложившихся растительных остатков, сохраняющих признаки анатомического строения. Горизонт Ehi – верхняя часть подзолистого горизонта, прокрашен потечным гумусом. Он имеет серовато-пепельный цвет (2.5Y 3/1 во влажном состоянии и 2.5Y 5/2 в сухом состоянии), что может быть связано с наличием мелких углистых частиц, а также потечного органического вещества, образующегося в процессе разложения подстилки. Диагностический подзолистый горизонт Е (2.5Y 5/1 во влажном состоянии и 2.5Y 7/2 в сухом состоянии) светлее и более однороден по цвету, чем вышележащий. Сменяющий его альфегумусовый горизонт разделен нами на два слоя – ВHF1 и ВHF2. ВHF1 имеет кофейно-охристый цвет (10YR 3/3 во влажном состоянии и 2.5Y 5/3 в сухом состоянии), супесчаный, уплотненный со слабовыраженной структурой. ВHF2 – палево-рыжий (10YR 4/3 во влажном состоянии и 2.5Y 6/1 в сухом состоянии), супесчаный, плотный, бесструктурный. Горизонт ВС – рыжевато-палевый (2.5Y 4/4 во влажном состоянии и 2.5Y 6/2 в сухом состоянии), плотный, мелкопесчаный, бесструктурный, постепенно переходит в почвообразующую породу. Горизонт Сg – палевый (2.5Y 6/1 во влажном состоянии и 2.5Y 6/3 в сухом состоянии) с сизым оттенком (2.5Y 5/1 во влажном состоянии и 2.5Y 6/1 в сухом состоянии) и охристыми пятнами (2.5Y 4/4 во влажном состоянии и 2.5Y 6/3 в сухом состоянии), плотный горизонт.

Рисунок 3. Профили серогумусовой метаморфизованной почвы и подзола иллювиально-железистого: cлева – контрольные профили, справа – профили траншей под валежными стволами P. abies: а – серогумусовая (дерновая) метаморфизованная почва со стволами 3, 4, 5а, 5б классов разложения; b – подзол иллювиально-железистый со стволами 3, 4, 5 классов разложения
Изменения морфологических свойств изучаемых почвенных объектов под влиянием валежных стволов P. abies 3-5 классов разложения проявляются в слабовыраженном изменении строения почвенных профилей, цвета срединных горизонтов и состава подгоризонтов подстилки (рис. 4). Процесс разложения валежного ствола P. abies длится приблизительно 100 лет (Ромашкин, 2021), а его влияние на морфологические свойства подстилающей почвы проявляется уже с 3 класса разложения (с 15-20 лет). В условиях серогумусовой почвы со стволами 3-5а классов разложения и в условиях подзола со стволами 3-4 классов изменения в строении профилей рассмотренных нами почвенных объектов прослеживаются непосредственно под валежем. В условиях серогумусовой (дерновой) метаморфизованной почвы со стволом 5б класса и подзола со стволом 5 класса разложения (с 35-40 лет) изменения в строении профиля наблюдаются на расстоянии до 25 см от валежного ствола.
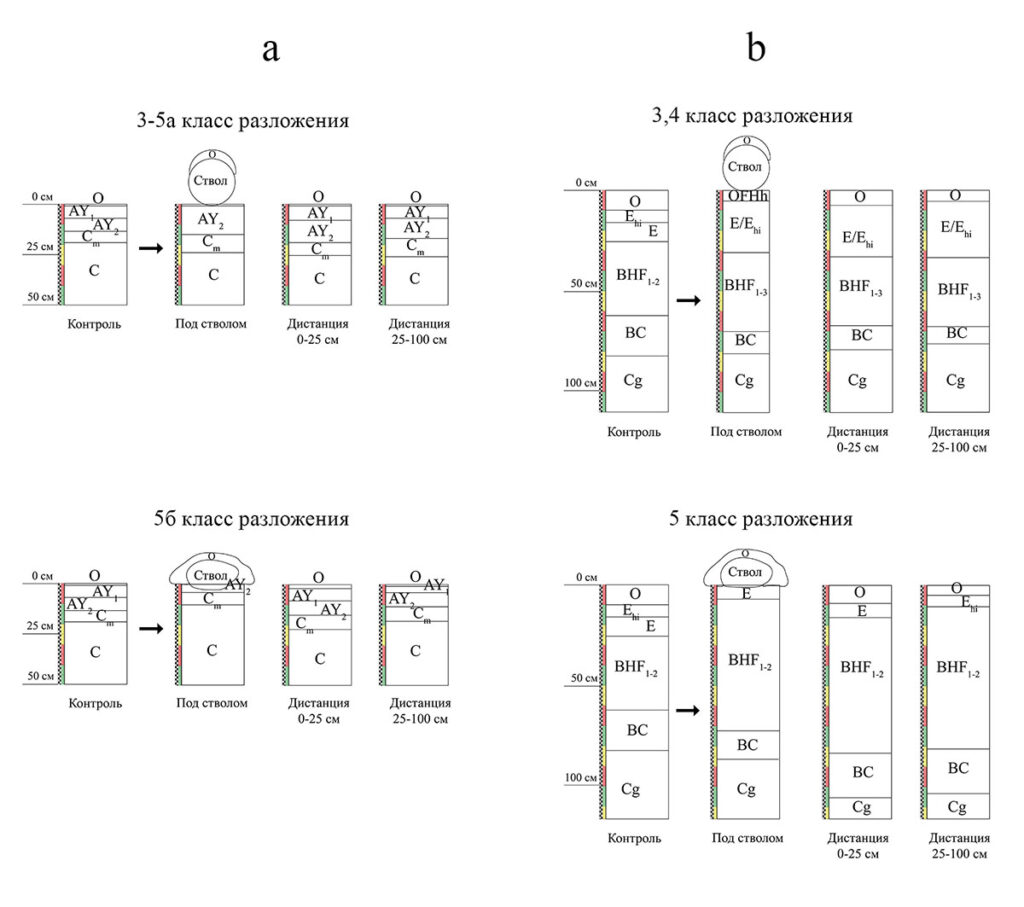
Рисунок 4. Схема изменения строения профилей почвенных объектов под валежным стволом P. abies 3-5 классов разложения и на расстоянии 0-25 и 25-100 см от ствола: a – в серогумусовой (дерновой) метаморфизованной почве со стволами 3-5а классов разложения и 5б класса разложения; b – в подзоле иллювиально-железистом со стволами 3-5 классов разложения
По мере разложения валежного ствола происходит его постепенное «встраивание» в почвенную толщу (рис. 4). Для валежных стволов 4 и 5 классов разложения характерно их частичное включение в верхние горизонты. В условиях серогумусовой (дерновой) метаморфизованной почвы происходит интегрирование преимущественно в горизонт AY, подзола иллювиально-железистого – в подстилку и горизонты E или Ehi, соответственно (рис. 4). Другими исследователями (Dhiedt et al., 2019) также было показано, что наиболее значимые изменения почвенных свойств происходят в поверхностных горизонтах. Это связано как с их непосредственным контактом с упавшим древесным стволом, так и c различными биогенно-аккумулятивными процессами (гумусообразование и гумусонакопление), способствующими укоренению растительности. Соответствующее влияние на эти процессы может оказывать почвенная фауна и микробиота, основная часть которых сосредоточена в поверхностных горизонтах.
В условиях серогумусовой метаморфизованной почвы влияние разлагающейся древесины прослеживается в исчезновении горизонта AY1 (рис. 4) и наличии под валежным стволом только горизонта AY2. AY1 – гумусово-аккумулятивный горизонт, формирование которого связано с подстилочным опадом и деятельностью почвенной фауны. Дождевые черви и прочая почвенная биота не только повышают плодородие, но и способствуют перемешиванию органо-минеральной части почвы. Накоплению органического вещества и его закреплению в верхней части профиля способствует также высокое содержание физической глины в подстилающей материнской породе. Падение ствола дерева на поверхность почвы препятствует поступлению свежего опада и его включению в процессы почвообразования. Кроме того, вымывание из ствола продуктов распада может способствовать увеличению подвижности органических веществ. Таким образом, горизонт AY1 под стволами поздних классов разложения валежного ствола перестает формироваться, а его остатки активно вовлекаются в состав разлагающейся древесной массы.
В подзоле иллювиально-железистом влияние разлагающейся древесины выражается в отсутствии горизонта Ehi под валежным стволом 5-го класса разложения. Данный горизонт имеет оттенки 2.5 Y 4/1 во влажном и сухом состояниях и присутствует на расстоянии 25-100 см от валежа (рис. 4). Под стволом и на расстоянии до 25 см от него вместо подзолистого горизонта с потеками гумуса (Ehi) четко просматривается хорошо отбеленный подзолистый горизонт E с оттенками 2.5 Y 5/1 и 2.5 Y 7/2 во влажном и в сухом состояниях соответственно. Похожие результаты были получены и для горизонта albic (подзолистого) в гумусово-железистых подзолах – Humo-Ferric Podzols (название по: The Canadian system …, 1998) в Британской Колумбии (Klinka et al., 1995). Более светлые оттенки горизонта E и отсутствие прокраски гумусом могут быть обусловлены отбеливанием минеральных зерен вследствие вымывания гумусовых веществ в нижележащие горизонты. Под валежным стволом фиксируется снижение величины pH, что обусловлено поступлением из валежной древесины более кислых продуктов разложения органических остатков (Spears, Lajtha, 2004; Krzyszowska-Waitkus et al., 2006). Содержание фульвокислот вблизи валежа выше по сравнению с таковым на начальных стадиях разложения (Zalamea, 2007). Фульвокислоты способствуют разрушению полевых шпатов, слоистых силикатов группы слюд, глинистых минералов и других минералов, находящиеся в почвах, что способствует перемещению продуктов разложения в нижние горизонты. Помимо этого, в условиях сильноразложившегося валежа происходит стимуляция биохимической активности (Wojciech, 2019).
Под валежными стволами подзола 3 и 4 классов разложения диагностируется подгоризонт OFHh (рис. 4) мощностью до 5 см. Он частично состоит из гумифицированного и хорошо мажущегося вещества. Отсутствие OFHh в контрольном почвенном разрезе и под стволом 5 класса разложения (рис. 4) позволяет предположить, что он формируется либо непосредственно в процессе деструкции валежного ствола из древесных остатков, либо он является нижней частью исходной редуцированной подстилки, в которой в процессе разложения древесины ствола появляются признаки перегнойности. Отсутствие данного подгоризонта под стволом 5 класса разложения может быть связано с интенсификацией почвенных процессов на последних стадиях деструкции.
Полученные нами данные о влиянии валежа на почвенные объекты на макроморфологическом уровне согласуются с результатами ранее проведенных исследований, свидетельствующими о способности валежа усиливать процессы элювиирования и оподзоливания (Krajina, Brooke 1965; Klinka et al., 1990). Тем не менее, некоторые из ранее описанных особенностей нами в настоящей работе не были отмечены. В частности, в исследованных нами типах почв не наблюдали: формирование более мощных и глубоких языковатых горизонтов albic под валежем (Wilding et al., 1983), увеличение мощности горизонта Ae под валежным стволом (Kayahara et al., 1996), лучшее развитие темноокрашенного горизонта spodic под валежной древесиной вследствие повышенных концентраций железа и алюминия в органических комплексах (Klinka et al., 1995). Отсутствие указанных признаков в рассмотренном нами подзоле иллювиально-железистом может быть связано с достаточно сильной щебнистостью его верхних горизонтов, препятствующей полноценному развитию верхних почвенных горизонтов.
На поверхности валежных стволов 3-5 классов разложения подзола иллювиально-железистого и серогумусовой метаморфизованной почвы происходит формирование новой подстилки (OL–OF) за счет отмирания растительности и накопления опада (рис. 4). В условиях подзола подстилка на поверхности валежа более четко дифференцирована на подгоризонты OL и OF, что может быть связано со спецификой условий их формирования – микрорельефом (микроповышение) и зарастанием валежа эпиксильной растительностью. В процессе разложения валежного ствола и, как следствие, исчезновения его контура горизонт O полностью встраивается в лесную подстилку.
ЗАКЛЮЧЕНИЕ
На макроморфологическом уровне валеж P. abies 3-5 классов разложения оказывает влияние на формирование верхних горизонтов почвенных объектов – подзола иллювиально-железистого и серогумусовой (дерновой) метаморфизованной почвы. Влияние валежного ствола прослеживается непосредственно под ним и на некотором расстоянии от него. В условиях серогумусовой почвы под стволами 3-5 классов разложения отсутствует горизонт AY1. В условиях подзола под валежом 3 и 4 классов разложения диагностируется подгоризонт OFHh, а также отсутствуют потеки гумуса в горизонте E под стволом 5-го класса разложения, что может свидетельствовать об интенсификации процессов элювиирования. На поверхности валежных стволов 3-5 классов разложения происходит формирование новой подстилки, имеющей строение OL–OF. В условиях серогумусовой (дерновой) метаморфизованной почвы на расстоянии до 25 см от ствола 5б класса разложения формируется такой же тип подстилки с ее дифференциацией на подгоризонты OL и OF. В профиле подзола влияние валежа выражается на расстоянии до 25 см так же, как и под стволом 5 класса разложения – более интенсивными процессами элювиирования.
БЛАГОДАРНОСТИ
Авторы выражают благодарность Ахметовой Гульнаре Вялитовне, старшему научному сотруднику лаборатории лесного почвоведения Института леса КарНЦ РАН, за помощь в диагностике почв. Авторы заявляют, что у них нет конфликта интересов. Финансовое обеспечение исследований осуществлялось из средств федерального бюджета на выполнение государственного задания КарНЦ РАН (Институт леса, 121061500082-2).
СПИСОК ЛИТЕРАТУРЫ
Васенев И. И. Влияние ветровала на развитие таежных подзолистых почв (на примере ельников центра Русской равнины): Дис. … канд. биол. наук, 1987. 329 с.
Васенев И. И. Почвенные сукцессии как форма эволюции почв таежных и антропогенно измененных лесостепных экосистем. М.: Изд-во МГУ, 2003. 49с.
Васенев И. И., Просвирина А. П. Влияние ветровальных нарушений на почвенный покров. Коренные темнохвойные леса южной тайги (резерват «Кологривский лес») / Под ред. А. В. Письмеровой. М.: Наука, 1988. С. 184–197.
Васенев И. И., Таргульян В. О. Ветровал и таежное почвообразование (режимы, процессы, морфогенез почвенных сукцессий). М.: Наука, 1995. 246 с.
Герасимова М. И., Чертов О. Г., Надпорожс-кая М. А. Формы гумуса в почвенных классификациях // Лесные почвы и функционирование лесных экосистем. Материалы VIII Всероссийской научной конференции с международным участием. М.: ЦЭПЛ РАН, 2019. С. 18–20.
Козлов М. В. Мнимые повторности (pseudoreplication) в экологических исследованиях: проблема, не замеченная российскими учеными // Журнал общей биологии. 2003. Т. 64. № 4. С. 292–307.
Острикова К. Т. Полевой определитель почв России. М.: Почвенный институт им. В. В. Докучаева, 2008. 182 c.
Радюкина А. Ю. Влияние валежа на свойства дерново-подзолистых почв // Лесоведение. 2004. №. 4. С. 51–60.
Ромашкин И. В. Динамика биогенных элементов в процессе разложения валежа в среднетаежных ельниках. Дис. … канд. биол. наук (спец. 1.5.15). Петрозаводск, 2021.167 с.
Самсонова В. П., Мешалкина Ю. Л. Часто встречающиеся неточности и ошибки применения статистических методов в почвоведении // Бюллетень Почвенного института им. В. В. Докучаева. 2020. № 102. С. 164–182.
Стороженко В. Г. Древесный отпад в коренных лесах Русской равнины. М.: Товарищество научных изданий КМК, 2011. 122 с.
Ханина Л. Г., Бобровский М. В., Смирнов В. Э., Иващенко К. В., Журавлева А. И., Романов М. С. Малонарушенные широколиственные леса: углерод и азот в валеже и почвах на массовых ветровалах // Научные основы устойчивого управления лесами. Материалы VIII Всероссийской научной конференции с международным участием. М.: ЦЭПЛ РАН, 2022. С. 130–133.
Швиденко А. З., Щепащенко Д. Г., Нильссон С. Оценка запасов древесного детрита в лесах России // Лесная таксация и лесоустройство. 2009. Вып. 1(41). С. 133–147.
Шишов Л. Л., Тонконогов В. Д., Лебедева И. И., Герасимова М. И. Классификация и диагностика почв России. М.: Ойкумена, 2004. 342 с.
Шорохова Е. В. Запасы и экосистемные функции крупных древесных остатков в таежных лесах. Дис. … докт. биол. наук (спец. 03.02.08). Санкт-Петербург, 2020. 299 с.
Яшин И. М., Атенбеков Р. А., Черников В. А., Васенев И. И. Экологическая роль водорастворимых органических веществ в гумусообразовании и миграции в почвах таежно-лесной зоны // Известия Тимирязевской сельскохозяйственной академии. 2018. №. 4. С. 32–45.
Błońska E., Lasota J. How decaying wood affects the accumulation of polycyclic aromatic hydrocarbons in soil of temperate mountain forest // Environmental Research. 2023. Vol. 223. Article number 115487, DOI: 10.1016/j.envres.2023.115487
Caron M. N., Kneeshaw D. D., De Grandpré L., Kauhanen H., Kuuluvainen T. Canopy gap characteristics and disturbance dynamics in old-growth Picea abies stands in northern Fennoscandia: Is the forest in quasi-equilibrium? // Annales Botanici Fennici. Finnish Zoological and Botanical Publishing Board. 2009. Vol. 46. No. 4. P. 251–262. DOI: 10.5735/085.046.0402
Dhar A., Forsch K. B. C., Naeth M. A. Effects of coarse woody debris on soil temperature and water content in two reconstructed soils in reclaimed boreal forest // Soil Systems. 2022. Vol. 6. No. 3. 62 p. DOI: 10.3390/soilsystems 6030062
Dhiedt E., De Keersmaeker L., Vandekerkhove K., Verheyen K. Effects of decomposing beech (Fagus sylvatica) logs on the chemistry of acidified sand and loam soils in two forest reserves in Flanders (northern Belgium) // Forest Ecology and Management. 2019. Vol. 445. P. 70–81. DOI: 10.1016/j.foreco.2019.05.006
Edman M., Jönsson M., Jonsson B. G. Fungi and wind strongly influence the temporal availability of logs in an old-growth spruce forest // Ecological Applications. 2007. Vol. 17. No. 2. P. 482–490. DOI: 10.1890/06-0852
Fraver S., Jonsson B. G., Jönsson M., Esseen P. A. Demographics and disturbance history of a boreal old-growth Picea abies forest // Journal of Vegetation Science. 2008. Vol. 19. No. 6. P. 789–798. DOI: 10.3170/2008-8-18449
Gonzalez-Polo M., Fernández-Souto A., Austin A. T. Coarse woody debris stimulates soil enzymatic activity and litter decomposition in an old-growth temperate forest of Patagonia, Argentina // Ecosystems. 2013. Vol. 16. P. 1025–1038. DOI: 10.1007/s10021-013-9665-0
Green R. N., Trowbridge R. L., Klinka K. Towards a taxonomic classification of humus forms // Forest Science. 1993. No. 39 (suppl. 1). P. a0001-z0002. DOI: 10.1093/forestscience/39.s1.a0001
Hagemann U., Moroni M.T., Gleißner J., Makeschin F. Accumulation and preservation of dead wood upon burial by bryophytes // Ecosystems. 2010. Vol. 13. No. 4. P. 600–611. DOI: 10.1007/s10021-010-9343-4
Harmon M. E., Franklin J. F., Swanson F. J., Sollins P., Gregory S. V., Lattin J. D., Cummins K. W. Ecology of coarse woody debris in temperate ecosystems // Advances in Ecological Research. 1986. Vol. 15. P. 133–302. DOI: 10.1016/ S0065-2504(08)60121-X
Hytteborn H., Verwijst T. The importance of gaps and dwarf trees in the regeneration of Swedish spruce forests: the origin and content of Sernander’s (1936) gap dynamics theory // Scandinavian Journal of Forest Research. 2011. Vol. 26. No. S10. P. 3–16. DOI: 10.1080/02827581.2011.517945
Jabiol B., Zanella A., Ponge J. F., Sartori G., Englisch M., Van Delft B., Le Bayon R. C. A proposal for including humus forms in the World Reference Base for Soil Resources (WRB-FAO) // Geoderma. 2013. Vol. 192. P. 286–294. DOI: 10.1016/j.geoderma.2011.05.016
Kayahara G. J., Klinka K., Lavkulich L. M. Effects of decaying wood on eluviation, podzolization, acidification, and nutrition in soils with different moisture regimes // Environmental Monitoring and Assessment. 1996. Vol. 39. P. 485–492. DOI: 10.1007/BF00396163
Khan K., Hussain A., Jamil M. A., Duan W., Chen L., Khan A. Alteration in Forest Soil Biogeochemistry through Coarse Wood Debris in Northeast China // Forests. 2022. Vol. 13. No. 11. Article number 1861. DOI: 10.3390/f13111861
Klinka K., Lavkulich L. M., Wang Q., Feller M. C. Influence of decaying wood on chemical properties of forest floors and surface mineral soils: a pilot study // Annales des Sciences Forestières. EDP Sciences. 1995. Vol. 52. No. 6. P. 523–533. DOI: 10.1051/forest:19950601
Klinka K., Wang Q., Carter R. E. Relationships among humus forms, forest floor nutrient properties, and understory vegetation // Forest Science. 1990. Vol. 36. No. 3. P. 564–581
Krajina V. J., Brooke R. C. Ecology of Western North America. Department of Botany, University of British Columbia Press, 1965.17 p.
Krzyszowska-Waitkus A., Vance G. F., Preston C. M. Influence of coarse wood and fine litter on forest organic matter composition // Canadian Journal of Soil Science. 2006. Vol. 86. No. 1. P. 35–46. DOI: 10.4141/S05-040
Kuuluvainen T., Wallenius T. H., Kauhanen H., Aakala T., Mikkola K., Demidova N., Ogibin B. Episodic, patchy disturbances characterize an old-growth Picea abies dominated forest landscape in northeastern Europe // Forest Ecology and Management. 2014. Vol. 320. P. 96–103. DOI: 10.1016/j.foreco.2014.02.024
Ma Y., Filley T. R., Szlavecz K., McCormick M. K. Controls on wood and leaf litter incorporation into soil fractions in forests at different successional stages // Soil Biology and Biochemistry. 2014. Vol. 69. P. 212–222. DOI: 10.1016/j.soilbio.2013.10.043
Moroni M. T., Hagemann U., Beilmann D. Deadwood is buried and preserved in a Labrador boreal forest // Ecosystems. 2010. Vol. 13. P. 452–458. DOI: 10.1007/s10021-010-9331-8
Munsell A. H. Munsell Soil Color Charts. Gretagmacbeth. 2000. 63 p.
Nazari M., Pausch J., Bickel S., Bilyera N., Rashtbari M., Razavi B. S., Zarebanadkouki M. Keeping thinning-derived deadwood logs on forest floor improves soil organic carbon, microbial biomass, and enzyme activity in a temperate spruce forest // European Journal of Forest Research. 2023. Vol. 142. No. 2. P. 287–300. DOI: 10.1007/s10342-022-01522-z
Piaszczyk W., Lasota J., Błońska E. Effect of organic matter released from deadwood at different decomposition stages on physical properties of forest soil // Forests. 2019. Vol. 11. No. 1. 24 p. DOI: 10.3390/f11010024
Schaetzl R. J., Follmer L. R. Longevity of treethrow microtopography: implications for mass wasting // Geomorphology. 1990. Vol. 3. P. 113–123. DOI: 10.1016/0169-555X(90)90040-W
Shannon V. L., Vanguelova E. I., Morison J. I. L., Shaw L. J., Clark J. M. The contribution of deadwood to soil carbon dynamics
in contrasting temperate forest ecosystems // European Journal of Forest Research. 2022. Vol. 141. No. 2. P. 241–252. DOI: 10.1007/s10342-021-01435-3
Shi B., Wang X., Yang S., Chen H., Zhao Y., Shen J., Huang B. Changes and driving factors of microbial community composition and functional groups during the decomposition of Pinus massoniana deadwood // Ecology and Evolution. 2024. Vol. 14. No. 4. Article number e11210, DOI: 10.1002/ece3.11210
Shorohova E., Kapitsa E. Stand and landscape scale variability in the amount and diversity of coarse woody debris in primeval European boreal forests // Forest Ecology and Management. 2015. Vol. 356. P. 273–284. DOI: 10.1016/j.foreco.2015.07.005
Sokołowski K., Gawryś R., Błońska E. The falling of a tree in the forest is the beginning of significant changes in the soil // Plant and Soil. 2024. Vol. 507. P. 383–396. DOI: 10.1007/s11104-024-06737-0
Spears J. D. H., Lajtha K. The imprint of coarse woody debris on soil chemistry in the western Oregon Cascades // Biogeochemistry. 2004. Vol. 71. P. 163–175. DOI: 10.1007/s10533-004-6395-6
Spears J. D., Holub S. M., Harmon M. E., Lajtha K. The influence of decomposing logs on soil biology and nutrient cycling in an old-growth mixed coniferous forest in Oregon, USA // Canadian Journal of Forest Research. 2003. Vol. 33. No. 11. P. 2193–2201. DOI: 10.1139/x03-148
Stutz K. P., Kaiser K., Wambsganss J., Santos F., Berhe A. A., Lang F. Lignin from white-rotted European beech deadwood and soil functions // Biogeochemistry. 2019. Vol. 145. No. 1-2. P. 81–105. DOI: 10.1007/s10533-019-00593-2
The Canadian System of Soil Classification. Soil Classification Working Group. Canadian Agricultural Services Coordinating Committee, Canada, Agriculture and Agri-Food Canada, Research Branch. NRC Research Press,1998. No. 1646. 187 p.
Ulanova N. G. The effects of windthrow on forests at different spatial scales: a review // Forest Ecology and Management. 2000. Vol. 135. No. 1-3. P. 155–167. DOI: 10.1016/S0378-1127(00)00307-8
Ulyshen M. D., Muller J., Seibold S. Bark coverage and insects influence wood decomposition: Direct and indirect effects // Applied Soil Ecology. 2016. Vol. 105. P. 25–30. DOI: 10.1016/j.apsoil. 2016.03.017
Wijas B. J., Cornwell W. K., Oberle B., Powel J. R., Zanne A. E. Faster than expected: release of nitrogen and phosphorus from decomposing woody litter // New Phytologist. 2025. Vol. 245. No. 5. P. 2214–2223. DOI: 10.1111/nph.20362
Wilding L. P., Smeck N. E., Hall G. F. Pedogenesis and Soil Taxonomy: the Soil Orders. Amsterdam; New York: Elsevier, 1983. 328 p.
Wojciech P., Ewa B., Jarosław L. Soil biochemical properties and stabilisation of soil organic matter in relation to deadwood of different species // FEMS microbiology ecology. 2019. Vol. 95. No. 3. Article number fiz011. DOI: 10.1093/femsec/fiz011
World Reference Base for Soil Resources. IUSS Working Group WRB. 2014. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports. FAO. Rome. 2014. No. 106. 189 p.
Zalamea M., González G., Ping C. L., Michaelson G. Soil organic matter dynamics under decaying wood in a subtropical wet forest: effect of tree species and decay stage // Plant and Soil. 2007. Vol. 296. P. 173–185. DOI: 10.1007/s11104-007-9307-4
Zanella A., Jabiol B., Ponge J. F., Sartori G., De Waal R., Van Delft B., … & Langohr R. European Humus Forms Reference Base. Elsevier, 2011. 56 p.
Рецензент: к. б. н., доцент Лаптева Е. М.





