- DOI 10.31509/2658-607x-2019-2-4-1-27
- УДК 630
Анализ факторов, влияющих на возобновление основных лесообразующих пород в сосновых лесах южного Подмосковья
![]() © 2019 г.
© 2019 г.
Г.Г. Фролова1*, П.В. Фролов1, В.Н. Шанин1,2,3, Н.В. Иванова3
1Институт физико-химических и биологических проблем почвоведения РАН – обособленное подразделение Федерального исследовательского центра “Пущинский научный центр биологических исследований РАН”
Россия, Московская обл., 142290, Пущино, ул. Институтская, д. 2, корп. 2
2 Центр по проблемам экологии и продуктивности лесов РАН
Россия, 117997, Москва, ул. Профсоюзная, 84/32, стр. 14
3 Институт математических проблем биологии РАН – филиал Федерального государственного учреждения «Федеральный исследовательский центр Институт прикладной математики им. М.В. Келдыша Российской академии наук»
Россия, Московская обл., 142290, Пущино, ул. проф. Виткевича, д. 1
*E-mail: gulfina.frolova@gmail.com
Поступила в редакцию 30.08.2019
Проведен количественный анализ зависимости жизненности и видового состава древесного подроста в сосновых лесах южного Подмосковья от комплекса биотических, абиотических и эдафических факторов. На каждом участке парных пробных площадей (с подростом и без него) были определены характеристики древостоя, доминанты живого напочвенного покрова, измерена освещенность, мощность подстилки и ее влажность, проведено определение концентрации углерода и азота в подстилке и органо-минеральном горизонте почвы. При обработке данных использовался анализ главных компонент, попарное сравнение экологических факторов на участках с подростом и без него, а также многомерный многофакторный корреляционный анализ. Результаты исследования показали, что основными факторами, влияющими на естественное лесовозобновление, являются характеристики богатства почвы и освещенность, в то время как зависимости возобновления деревьев от доминантов живого напочвенного покрова не выявлено.
Ключевые слова: древесный подрост, естественное лесовозобновление, экологические факторы, живой напочвенный покров.
Важность изучения функционирования лесных экосистем определяется их глобальным значением для биосферы (Демаков, 2000; Уткин и др., 2001). Важнейшим показателем устойчивого развития лесов, особенно в условиях, далеких от экологического оптимума, является успешное естественное возобновление древостоя (Денисов и др., 2009). Данный процесс продолжителен по времени (Восточноевропейские…, 2004) и зависит от множества взаимодействующих факторов. Так, Г.Ф. Морозов (1912) выделяет 5 групп факторов, влияющих на естественное лесовозобновление: (1) биологические и экологические; (2) лесорастительные условия; (3) ценотические; (4) антропогенные; (5) историко-географические. Среди множества факторов можно выделить основные, влияющие на формирование подроста.
Факторы, влияющие на прорастание семян. Наличие семян на поверхности почвы в большей степени зависит от близости к генеративным деревьям. Большая часть упавших семян сосны (Pinus sylvestris L.) располагается в пределах 18–20 м от родительского дерева (Booth, 1984). Менее 2% семян ели (Picea abies (L.) H.Karst.), P. sylvestris и березы (Betula spp.) прорастают в естественных условиях, при этом всхожесть семян прямо пропорциональна их размеру (Yazdani, Lindgren, 1990; Beland et al., 2000). Высота и плотность подроста отрицательно коррелируют с близостью деревьев верхних ярусов в сосновых лесах (Siipilehto, 2006), поскольку конкуренция за ресурсы препятствует развитию подроста (Montes, Canellas, 2007; Ruuska et al., 2008).
Влияние освещенности на выживание подроста. Освещенность под пологом является одним из ведущих факторов, влияющих на выживание проросших семян Betula spp., P. sylvestris и Quercus robur L. Было обнаружено, что семена могут прорастать при слабом освещении, но их выживание зависит от наличия достаточного количества света (Кобак, Алексеев, 1965; Алексеев, 1975; Евстигнеев, 2012; Gill, 1992; Nilsson et al., 1996; Poorter, De Jong, 1999; Poorter et al., 2012; Wyka et al., 2012). Наблюдается заметное снижение прироста биомассы и отношения массы подземных органов к массе надземных при низкой интенсивности света, что может влиять на долгосрочную выживаемость проростков (Gong, 1992). При этом Кобан с соавторами (Coban et al., 2016) показали, что проростки P. sylvestris могут выживать в течение 10–12 лет при низкой освещенности, а формирование окон в пологе и увеличение освещенности может инициировать активный рост соснового подроста (Полякова, 1971; Рязапов, Кабанов, 2011). Подрост P. abies, являющейся позднесукцессионным, теневыносливым видом (Восточноевропейские…, 2004; Niinemets, Valladares, 2006), обычно проявляет еще более высокую устойчивость к затенению, чем взрослые деревья, что связано с особенностями соотношения масс органов, продуцирующих и потребляющих углеводы (Dyderski et al., 2018).
Влияние почвенного богатства. Известно, что успешность естественного возобновления сосны также зависит от локальных характеристик почвы (Wittich, 1955). Например, толстый слой подстилки может замедлить или даже подавить появление проростков, предотвращая контакт между семенами и минеральной почвой (Чучалина, Мищихина, 2013). В литературных источниках встречаются противоречивые данные о влиянии почвенного богатства на естественное возобновление P. abies. Так, в некоторых работах приводятся данные о том, что подрост P. abies встречается чаще в более бедных местообитаниях (Miina, Saksa, 2013). В то же время, в других исследованиях отмечается более активное возобновление ели в более богатых местообитаниях (Szydlarski, Modrzyński, 2015). Подобные различия, однако, могут быть связаны с климатическими факторами. Естественное возобновление Q. robur может происходить в различных по богатству почв условиях (Aas, 2002), но наиболее активно оно происходит на богатых суглинистых и глинистых почвах (Annighöfer et al., 2015).
Влияние влажности почвы. В большинстве работ отмечается приуроченность подроста P. abies и Q. robur к местообитаниям с высокой влажностью (Ellenberg, Leuschner, 2010; Gutowski, Jaroszewicz, 2015; Caudullo et al., 2016; Čater, Batič, 2006). Корневая конкуренция за воду и минеральное питание с деревьями верхних ярусов и растениями живого напочвенного покрова (ЖНП) может быть ведущим лимитирующим фактором, оказывающим влияние на процесс возобновления на разных его этапах (Грязькин, 2001; Цветков, 2009; Дружинин, 2007; Смирнов, 2007; Шаньгина, 2010; Kielland-Lund, 1981; Kuuluvainen, Ylläsjärvi, 2011; Axelsson et al., 2014).
Влияние напочвенного покрова. Выделяют 4 типа воздействий, которые оказывает ЖНП на лесовозобновительные процессы: механическое (например, плотный мохово-лишайниковый покров может препятствовать проникновению корней проростков в почву); физическое (формирование напочвенного микроклимата, неподходящего для развития проростков деревьев); физиологическое (растения создают конкуренцию за элементы питания, влагу); биохимическое (влияние, опосредованное выделением некоторыми видами ЖНП фитонцидов и колинов) (Денисов и др., 2009). Эти воздействия могут комбинироваться разным образом и в результате оказывать как положительный, так и отрицательный эффект на начальные этапы возобновления леса (Санников, Санникова, 1985).
Процессы, регулирующие динамику возобновления, и их взаимодействие, по своей сути сложны, и определить вклад конкретного фактора в общий процесс возобновления древостоя без комплексного мониторинга экологических условий под пологом не представляется возможным.
Целью данного исследования является выявление взаимосвязи жизненности и видового состава древесного подроста в сосновых лесах с комплексом экологических факторов.
Под жизненностью понимается степень процветания особи в ценозе и перспектив ее дальнейшего развития (Гатцук, Ермакова, 1987; Жукова, 2001). Среди факторов рассмотрены следующие: абиотические (освещенность под пологом), биотические (структура ЖНП) и эдафические (влажность почвы, мощность подстилки и верхнего минерального горизонта почвы, концентрация биофильных элементов в них).
МАТЕРИАЛЫ И МЕТОДЫ
Сосновые леса широко распространены в Подмосковье, их площадь составляет 427 тыс. га (Лесной план Московской области, 2018). В южном Подмосковье наиболее распространены сосняки кустарничково-зеленомошные, зеленомошные, вейниково-черничные и долгомошно-сфагновые (Суслова, 2019), произрастающие на дерново-слабо- и среднеподзолистых почвах супесчаного и песчаного гранулометрического состава, сформировавшихся на моренных и покровных суглинках и аллювиальных отложениях террас Оки (Экологический мониторинг, 1989). Для данных почв характерно пониженное содержание органического вещества, азота, основных катионов и большинства микроэлементов (Комаров и др., 2006). Полевые исследования были проведены в весенне-летний период 2018–2019 годов в сложных сосняках опытного лесничества «Русский лес» и Приокско-Террасного государственного природного биосферного заповедника (Серпуховский район Московской области), в составе которых представлены дуб черешчатый, липа сердцевидная (Tilia cordata Mill.), клен остролистный (Acer platanoides L.), береза, осина обыкновенная (Populus tremula L.) и ель европейская. Согласно материалам таксации 1999 г., средний возраст древостоя – 53 года; средняя полнота – 0.73; средний класс бонитета – 1.4. Подлесок представлен рябиной обыкновенной (Sorbus aucuparia L.), крушиной ломкой (Frangula alnus Mill.). Схема расположения ПП приведена на рисунке 1.
Рисунок 1. Схема расположения пробных площадей
Описание пробных площадей. Заложено 13 временных пробных площадей (ПП), каждая из которых состояла из двух смежных участков (табл. 1): на одном участке подрост присутствовал (учитывались особи видов деревьев семенного происхождения, начиная с ювенильного онтогенетического состояния, высотой до 1.5 м), а на прилегающем участке подроста не было. Пробные площади выбирались таким образом, чтобы высота древостоя визуально не различалась, и группа возраста соответствовала средневозрастным или приспевающим древостоям. Участок с наличием подроста (не менее 40 особей) учитывался как «опыт», а участок без подроста – как «контроль». Размер каждого участка составлял 10×10 м.
Таблица 1. Краткое описание пробных площадей
|
№ ПП |
Наличие подроста |
Формула древостоя |
Полнота древостоя, м2 га−1 |
Тип леса |
|
1 |
— |
6С4Б |
36.1 |
Сосняк голубичный сфагновый |
|
+ |
6Б4С |
27.8 |
Березняк кустарничково-осоковый сфагновый |
|
|
2 |
— |
9С1Б |
31.6 |
Сосняк черничный |
|
+ |
8С2Б |
30.7 |
Сосняк черничный |
|
|
3 |
— |
10С |
23.8 |
Сосняк чернично-орляковый |
|
+ |
10С+Б |
28 |
Сосняк черничный |
|
|
4 |
— |
9С1Б |
22 |
Сосняк зеленомошный |
|
+ |
10С |
25.8 |
Сосняк кустарничково-зеленомошный |
|
|
5 |
— |
10С |
24 |
Сосняк зеленомошный |
|
+ |
10С |
22.8 |
Сосняк зеленомошный |
|
|
6 |
— |
10С |
26 |
Сосняк зеленомошный |
|
+ |
10С |
16.8 |
Сосняк зеленомошный |
|
|
7 |
— |
10С |
19.1 |
Сосняк зеленомошный |
|
+ |
10С+Б |
20 |
Сосняк зеленомошный |
|
|
8 |
— |
6С4Б+Ос |
16.6 |
Сосняк чернично-вейниковый |
|
+ |
5С5Б+Ос |
26.2 |
Сосняк вейниковый |
|
|
9 |
— |
5С3Б2Е |
25.5 |
Сосняк мертвопокровный |
|
+ |
6С2Е2Б |
24.8 |
Сосняк мертвопокровный |
|
|
10 |
— |
10С |
26.5 |
Сосняк зеленомошный |
|
+ |
10С |
28 |
Сосняк зеленомошный |
|
|
11 |
— |
5Е3С2Б |
25.2 |
Ельник мертвопокровный |
|
+ |
6С3Е1Б |
23.2 |
Сосняк кустарничково-зеленомошный |
|
|
12 |
— |
7С2Е1Б+Д |
25.7 |
Сосняк вейниковый |
|
+ |
8С1Е1Д+Б |
25.3 |
Сосняк вейниковый |
|
|
13 |
— |
10С |
24.5 |
Сосняк зеленомошный |
|
+ |
10С |
21.8 |
Сосняк зеленомошный |
На каждом участке были определены абсолютная полнота и формула древостоя с помощью цепного реласкопа. В связи с отсутствием выраженной ярусности на большинстве ПП, полнота и формула древостоя отдельно по ярусам не выделялись. Были определены доминанты травяно-кустарничкового и мохово-лишайникового ярусов, измерена мощность подстилки (по 5 измерений на каждом участке) и ее влажность (по 60 измерений на каждом участке), произведен отбор образцов органогенного (подстилка) и органо-минерального горизонтов почвы для определения концентрации углерода и азота: по одной смешанной пробе (метод конверта) с каждого участка. Для определения освещенности на каждом участке была произведена съемка в зенит при помощи фотоаппарата Canon D600 с циркулярным полусферическим объективом Sigma AF 4.5/2.8 EX DC HSM Fisheye Canon, угол обзора которого равен 180 градусам (одна фотография выполнялась в центре участка и 4 – на расстоянии 1 м от углов участка по направлению к его центру). На участках с подростом была определена жизненность каждой особи подроста (3 балла – повышенная, 2 балла – нормальная и 1 балл – пониженная жизненность) в соответствие с методикой Г.Г. Жиляева (2005). Характеристика пробных площадей приведена в таблице 2.
Методика расчета освещенности. Фотографирование проводилось в зенит (выравнивание фотоаппарата проводилось при помощи карданова подвеса), верх кадра ориентировался на истинный север с учетом магнитного склонения, которое вычислялось для района съемки по всемирной модели геомагнитного поля Земли WMM (Chulliat et al., 2015).
Для выявления порогового значения для разделения пикселей, относящихся к небу и к древесному пологу, и, таким образом, получения бинарного изображения (Wagner, 1998; Jonckheere et al., 2004; Jonckheere et al., 2005; Cescatti, 2007) использовался синий канал цветовой модели RGB. Это связано с тем, что элементы листвы имеют гораздо меньшую отражающую способность и коэффициент пропускания в синей области видимого электромагнитного спектра (Macfarlane et al., 2000; Leblanc et al., 2005; Zhang et al., 2005), в то время как различие яркости облаков и голубого неба в данной области спектра минимально. В связи с тем, что пороговое значение, подобранное вручную, может являться источником ошибок из-за его субъективности (Rich, 1990; Jonckheere et al., 2004; Cescatti, 2007; Jarčuška et al., 2010), был применен автоматический метод вычисления порога бинаризации Нобиса-Хунцикера (Nobis, Hunziker, 2005), не зависящий от оператора.
Таблица 2. Характеристика пробных площадей. C – P. sylvestris, Е – P. abies, Д – Q. robur, Б – Betula spp.
|
№ ПП |
Наличие подроста |
Преобладающая жизненность подроста, С | Е | Д | Б |
GLI, %±SD (n=5) |
Об. влажность в слое 0–6 см, %±SD (n=60) |
Мощность подстилки, см±SD (n=5) |
Сорг подстилки, %±SD (n=5) |
Nобщ подстилки, %±SD (n=5) |
Сорг мин. горизонта, %±SD (n=5) |
Nобщ мин. горизонта, %±SD (n=5) |
|
1 |
— |
– | – | – | – |
23.59±10.38 |
19.43±5.18 |
10.00±4.54 |
41.93±5.60 |
43.37±6.03 |
1.45±0.18 |
1.54±0.24 |
|
+ |
2 | 3 | 2 | 2 |
23.20±7.12 |
20.84±9.53 |
9.80±2.67 |
32.38±3.78 |
25.16±2.64 |
1.19±0.17 |
0.86±0.12 |
|
|
2 |
— |
– | – | – | – |
19.21±7.79 |
26.07±10.94 |
6.60±1.38 |
37.11±6.44 |
5.78±0.93 |
1.41±0.16 |
0.24±0.04 |
|
+ |
2 | 3 | 2 | 2 |
25.60±12.05 |
9.31±3.68 |
11.80±2.92 |
38.18±4.48 |
8.99±1.58 |
1.47±0.25 |
0.31±0.05 |
|
|
3 |
— |
– | – | – | – |
19.49±9.15 |
17.50±7.39 |
6.20±1.47 |
40.29±7.58 |
3.84±0.74 |
1.41±0.24 |
0.16±0.02 |
|
+ |
1 | 3 | 3 | – |
25.00±8.57 |
10.61±3.18 |
7.80±1.87 |
38.51±3.86 |
5.47±0.88 |
1.33±0.16 |
0.21±0.03 |
|
|
4 |
— |
– | – | – | – |
14.59±7.21 |
28.73±11.51 |
11.20±4.94 |
39.51±6.98 |
5.46±0.88 |
1.29±0.18 |
0.20±0.03 |
|
+ |
1 | 3 | 2 | – |
29.52±12.55 |
11.48±3.77 |
6.60±2.84 |
41.78±7.79 |
5.16±0.67 |
1.44±0.18 |
0.19±0.03 |
|
|
5 |
— |
– | – | – | – |
6.50±2.64 |
25.68±9.04 |
3.80±1.66 |
32.54±5.60 |
8.21±0.91 |
1.09±0.19 |
0.32±0.05 |
|
+ |
– | – | 2 | – |
21.86±7.48 |
4.60±2.00 |
6.80±2.15 |
42.42±7.45 |
11.04±1.42 |
1.47±0.25 |
0.42±0.05 |
|
|
6 |
— |
– | – | – | – |
4.84±1.74 |
21.84±6.74 |
3.20±1.40 |
37.56±4.71 |
8.30±1.28 |
1.51±0.20 |
0.32±0.06 |
|
+ |
1 | – | 3 | 2 |
27.72±12.55 |
4.61±2.06 |
2.00±0.51 |
32.25±5.18 |
4.08±0.72 |
1.39±0.23 |
0.20±0.04 |
|
|
7 |
— |
– | – | – | – |
8.10±2.73 |
16.52±7.03 |
2.60±0.94 |
38.61±5.47 |
10.78±1.98 |
1.54±0.17 |
0.47±0.09 |
|
+ |
3 | 2 | 2 | 2 |
16.54±5.65 |
5.98±2.87 |
3.60±0.99 |
41.21±5.25 |
10.94±1.91 |
1.60±0.28 |
0.47±0.05 |
|
|
8 |
— |
– | – | – | – |
23.68±8.42 |
17.65±3.96 |
7.40±3.53 |
37.00±5.25 |
5.02±0.63 |
1.54±0.18 |
0.25±0.04 |
|
+ |
2 | 3 | 2 | – |
19.85±4.00 |
12.61±3.23 |
5.40±1.87 |
34.15±3.81 |
5.39±0.69 |
1.30±0.18 |
0.27±0.03 |
|
|
9 |
— |
– | – | – | – |
21.11±7.67 |
11.62±4.38 |
3.00±1.26 |
36.04±5.37 |
2.93±0.41 |
1.58±0.17 |
0.16±0.03 |
|
+ |
– | 2 | 2 | – |
10.59±2.36 |
17.31±6.49 |
1.80±0.83 |
32.86±5.71 |
3.98±0.77 |
1.41±0.21 |
0.22±0.04 |
|
|
10 |
— |
– | – | – | – |
9.20±3.56 |
21.45±5.72 |
2.60±0.69 |
30.46±3.44 |
6.26±0.69 |
1.15±0.13 |
0.28±0.05 |
|
+ |
2 | 3 | 3 | 2 |
21.71±8.44 |
7.67±2.27 |
3.80±1.64 |
20.86±3.88 |
3.31±0.59 |
0.95±0.17 |
0.17±0.02 |
|
|
11 |
— |
– | – | – | – |
14.42±3.82 |
9.47±1.95 |
4.80±1.44 |
36.92±6.12 |
5.91±1.17 |
1.52±0.28 |
0.23±0.04 |
|
+ |
– | 2 | 2 | – |
9.72±2.77 |
9.73±2.23 |
8.60±3.57 |
38.16±7.48 |
3.50±0.48 |
1.50±0.24 |
0.13±0.02 |
|
|
12 |
— |
– | – | – | – |
11.73±4.52 |
13.41±6.51 |
5.20±2.12 |
32.13±4.88 |
2.22±0.35 |
1.33±0.22 |
0.11±0.02 |
|
+ |
– | 2 | 2 | – |
8.06±3.56 |
7.61±3.54 |
8.40±2.41 |
34.27±6.84 |
2.42±0.30 |
1.58±0.22 |
0.12±0.02 |
|
|
13 |
— |
– | – | – | – |
7.05±2.96 |
27.78±8.37 |
5.50±1.25 |
40.97±5.38 |
5.18±0.73 |
1.61±0.18 |
0.20±0.02 |
|
+ |
2 | – | 2 | 2 |
25.73±8.72 |
6.66±2.82 |
6.40±2.55 |
36.13±7.15 |
3.64±0.58 |
1.53±0.16 |
0.14±0.03 |
Полученное черно-белое изображение использовалось в качестве маски для расчета того, какая доля поступающего света пропускается через эту маску, то есть определяет пропускание диффузного и прямого излучения. Световой режим под пологом определяется не только количеством и размером просветов в пологе, но и тем, насколько расположение их проекций совпадает с траекторией Солнца на небосклоне. Приходящее на полог излучение варьирует в соответствии с суточным и годовым циклом, а также зависит от условий облачности. Для вычисления надпологовой прямой и рассеянной радиации используются климатические данные, полученные из радиационной модели PVGIS (Šúri et al., 2005). Объединение этой модели с маской пропускания излучения (бинаризованным полусферическим изображением) позволяет прогнозировать мощность излучения, доступную растениям живого напочвенного покрова на конкретной территории за вегетационный период (установленный для изучаемой территории по датам перехода среднесуточной температуры через +5 градусов, со 102 по 289 день года (Николаев, Аблеева, 2015)) и средневзвешенный показатель пропускания излучения GLI. Для автоматизации обработки фотографий в соответствии с данным алгоритмом использовался программный продукт Hemisfer (Thimonier et al., 2010).
Вычисление объемной влажности почвы по показаниям влагомера. Для измерений влажности почвы использовался влагомер грунта МГ-44, предназначенный для измерения влажности почвы в слое 0–6 см при помощи чувствительного радиочастотного датчика. Принцип работы прибора основан на зависимости диэлектрических свойств пористой среды от ее влажности. Для измерений использовалась заводская градуировка «Абсолют» (дающая наиболее детализированную шкалу показаний: 0–100), показания которой рассматривались как промежуточные «условные единицы», с последующим пересчетом по дополнительной («корректирующей») кривой связи фактических значений объемной влажности исследуемой почвы с «условными» показаниями прибора по упомянутой шкале. «Корректирующая» кривая строилась по лабораторным измерениям. Измерения прибором МГ–44 проводились в 10-литровом пластиковом ведре, заполненном исследуемой почвой слоем 15 см (объем около 6 л). Объем и плотность определялись по высоте слоя (уровню) почвы в сосуде. Измерения прибором проводились в центральной части сосуда в 5-кратной повторности. После выполнения измерений к сухой почве добавляли определенную порцию воды (200 мл), пробу перемешивали, уплотняли, выдерживали 20 мин., производили отсчеты по прибору, и цикл «долив – перемешивание – выдержка – измерение» повторяли вплоть до полного насыщения почвы водой. После этого сосуд оставляли открытым, а измерения повторяли по мере высыхания почвы с интервалом от 2–3 дней в начале, до 1 недели в конце эксперимента, до достижения постоянного веса. Перед измерениями почву перемешивали (т.к. высыхание шло преимущественно с поверхности), уплотняли, выдерживали 20 мин. и взвешивали. Потерю влаги оценивали по потере массы.
В значительной части рассматриваемого диапазона объемной влажности различия между показаниями прибора для исследуемых почв оказались невелики, что допускает использование «универсальной» кривой, с помощью которой был проведен пересчет показаний влагомера в объемную влажность:
𝑦 = (−62.57 ×𝑥) / ( 𝑥 −187.2)
где y – объемная влажность, x – показания влагомера, R2 = 0.97.
Обработка почвенных проб. Первичные почвенные пробы высушивали в сушильном шкафу при температуре 90 ºC до абсолютно сухого состояния. Достижение постоянной массы образцов контролировали посредством взвешивания на электронных лабораторных весах с точностью до 0.001 г. Постоянная масса образца считалась достигнутой, если разность трех последовательных измерений не превышала погрешности взвешивания. Высушенные пробы были уменьшены квартованием (делением пробы по диагоналям на 4 части). Две противоположно расположенные части были использованы в качестве средней лабораторной пробы. Среднюю лабораторную почвенную пробу разделяли на квадраты, и из каждого квадрата шпателем отбирали небольшое количество почвы. Полученную таким образом аналитическую пробу измельчали в ступке и просеивали через сито с отверстиями диаметром 0.25 мм. Измельчение и просеивание осуществлялось до тех пор, пока все почвенные частицы не прошли сквозь сито. В подготовленных пробах проводили измерения концентраций углерода и азота при помощи CHN анализатора CHNS-932 фирмы LECO (США).
Анализ полевых данных. Типы сообществ определяли по доминанту ЖНП. Анализировали видовое разнообразие древесного подроста в сообществах разных типов.
Для выделения основных факторов, влияющих на успешность возобновления, использовали анализ главных компонент (PCA). Непрямую ординацию участков по физико-химическим характеристикам почвы, ее влажности, а также оценке освещенности исследованных древостоев выполняли с помощью функции prcomp пакета stats в среде статистического программирования R (R Core Team, 2019). Интерпретацию осей ординации давали на основе значений коэффициентов корреляции с рассматриваемыми факторами.
Затем проводили попарные сравнения объемной влажности органогенных горизонтов, GLI, мощности подстилки, соотношения C:N в органогенных и органоминеральных горизонтах в парах «контроль – опыт». Для этого выполняли проверку статистической гипотезы о равенстве средних значений двух выборок. Для проверки гипотезы о равенстве средних использовали парный критерий Стьюдента (для выборок, распределение которых соответствовало нормальному распределению) и парный критерий Вилкоксона (для выборок, распределение которых не соответствовало нормальному распределению). Предварительно была проведена проверка всех выборок на нормальность распределения при помощи критерия Шапиро-Вилка при уровне значимости 0.05.
Для выявления взаимосвязей между анализируемыми факторами использовался многомерный многофакторный корреляционный анализ с использованием коэффициента корреляции Пирсона.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Влияние типа сообщества. Исследованные площадки были объединены в группы, выделенные по доминантам ЖНП: зеленомошную (5 площадок), черничную (2 площадки), кустарничково-зеленомошную (2 площадки), вейниковую (2 площадки), кустарничково-осоковую сфагновую (1 площадка) и мертвопокровную (1 площадка). В связи с тем, что число площадок в каждой группе недостаточно для количественного анализа разнообразия подроста, его оценка была проведена на качественном уровне. Видовой состав подроста различался между выделенными типами сообществ (рис. 2). Наибольшим видовым разнообразием (4 вида) характеризовались черничные, кустарничково-осоковые сфагновые и зеленомошные леса. В кустарниково-зеленомошных и вейниковых (из Calamagrostis arundinacea Roth) сообществах отсутствовал подрост березы. В мертвопокровных лесах в подросте отмечено два вида, при этом выше была доля P. abies, участие дуба было незначительным.
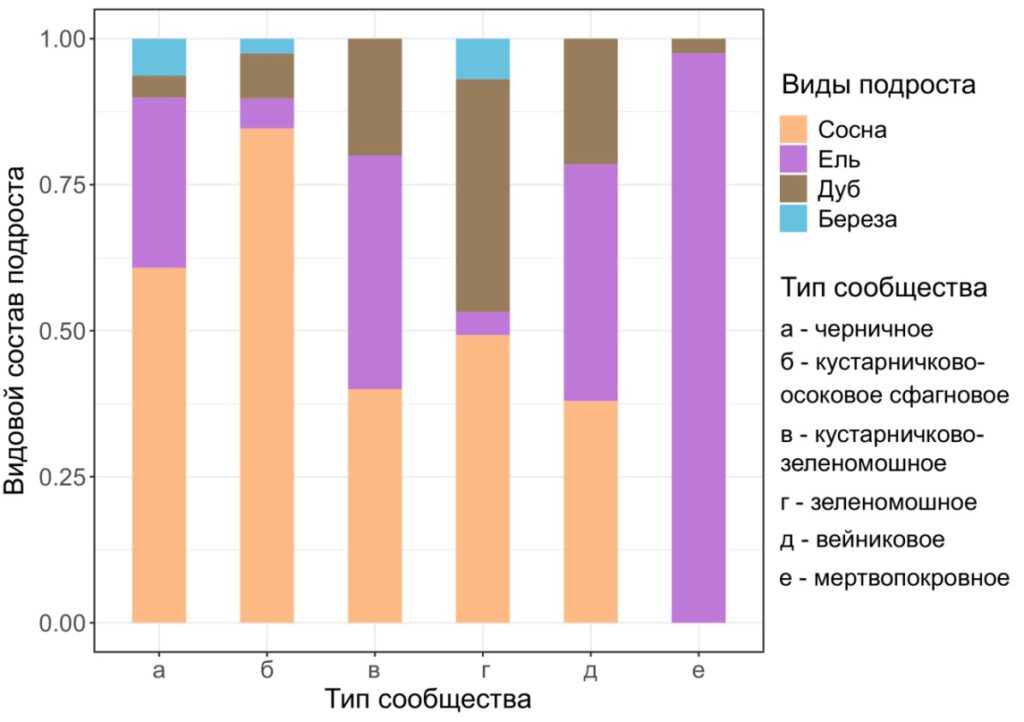
Рисунок 2. Видовой состав подроста в разных типах исследованных сообществ
Однако отметим, что зачастую экологические условия и корневая конкуренция с растениями верхнего яруса оказывают более существенное влияние на выживание древесного подроста, чем конкуренция с растениями ЖНП (Санников, Санникова, 2014).
Влияние экологических факторов на наличие подроста. Проведенный анализ главных компонент показал, что первые две оси объясняют 76.5% вариации между площадками с присутствием подроста и без него (рис. 3). Наибольший вклад в 1-ю ось вносят факторы, определяющие почвенное богатство (соотношение C:N в органогенных и органоминеральных горизонтах почвы, а также мощность подстилки), во 2-ю ось – освещенность под пологом и влажность почвы в слое 0–6 см.
Участки с подростом расположены в нижней части ординационной диаграммы, в области более высоких значений освещенности и меньшей влажности. При этом характер зависимости доли подроста от рассматриваемых факторов различался для разных видов. Для сосны определяющими являлись освещенность и влажность: высокая доля подроста P. sylvestris отмечена на хорошо освещенных сухих участках. Наибольшая доля подроста ели наблюдалась на участках с высокими показателями почвенного богатства. Участки с высокой долей дубового и березового подроста характеризовались высокой освещенностью и низкой влажностью, а также высокими показателями почвенного богатства. Основными факторами, влияющими на жизненность подроста, были освещенность и влажность. Подрост всех рассматриваемых видов характеризовался более высокой жизненностью при более высоких значениях GLI и низкой влажности.
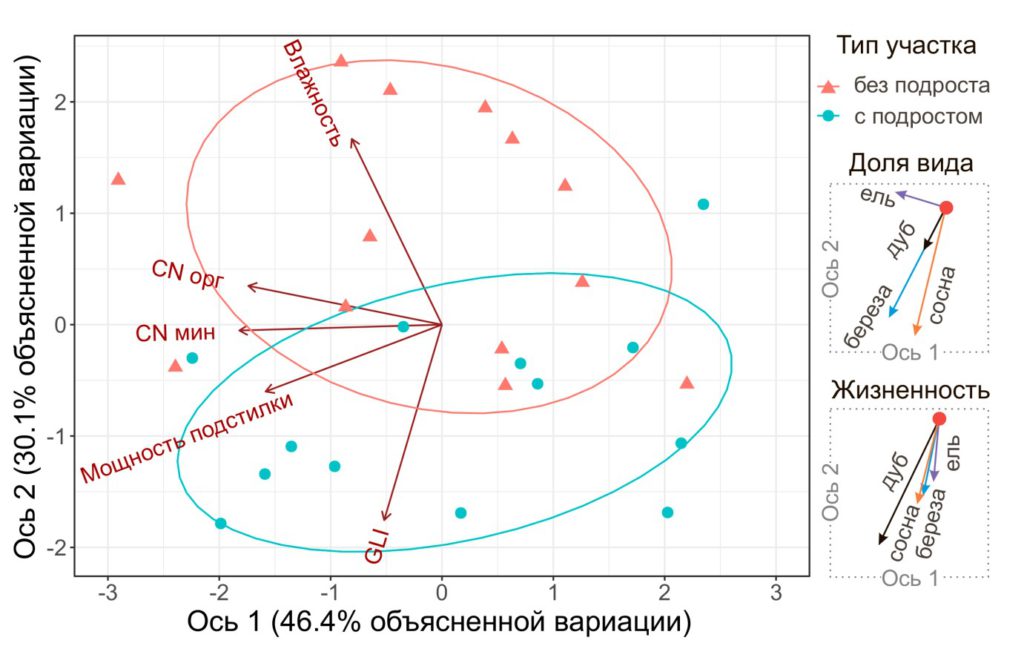
Рисунок 3. Результаты ординации исследованных участков методом анализа главных компонент (PCA)
Таким образом, результаты ординационного анализа согласуются с выявленными ранее (Evstigneev, 2018) видоспецифичными особенностями древесного подроста. Для возобновления светолюбивой сосны принципиальным является высокий уровень освещенности, и, в то же время, ее подрост способен выживать в местообитаниях с бедными почвами (табл. 2). Напротив, основным фактором успешного возобновления теневыносливой ели является почвенное богатство, тогда как освещенность не оказывала влияния в пределах изученного диапазона (рис. 3). Береза и дуб, согласно ординации, занимают промежуточное положение, но освещенность и влажность оказывают большее влияние, по сравнению с почвенным богатством. Ввиду очевидной зависимости влажности подстилки от инсоляции можно считать уровень освещенности фактором, определяющим наличие подроста большинства видов деревьев. Также об этом может свидетельствовать выявленная нами зависимость жизненности подроста только от влажности почв и освещенности. Однако следует отметить, что в рамках данной работы мы изучали факторы, влияющие на жизненность подроста, в то время как на вероятность заноса и прорастания семян могут влиять другие факторы (например, возникновение нарушений ЖНП).
Экологические факторы на пробных площадях. Средние значения GLI, мощности подстилки, а также соотношения C:N органо-минерального горизонта, как правило, были выше на площадках с подростом, а средние значения объемной влажности почвы в слое 0–6 см и соотношения C:N подстилки на них были ниже (рис. 4). При этом попарное сравнение экологических условий в парах «контроль – опыт» показало наличие статистически значимых различий только для объемной влажности почвы в слое 0–6 см (p = 0.002).
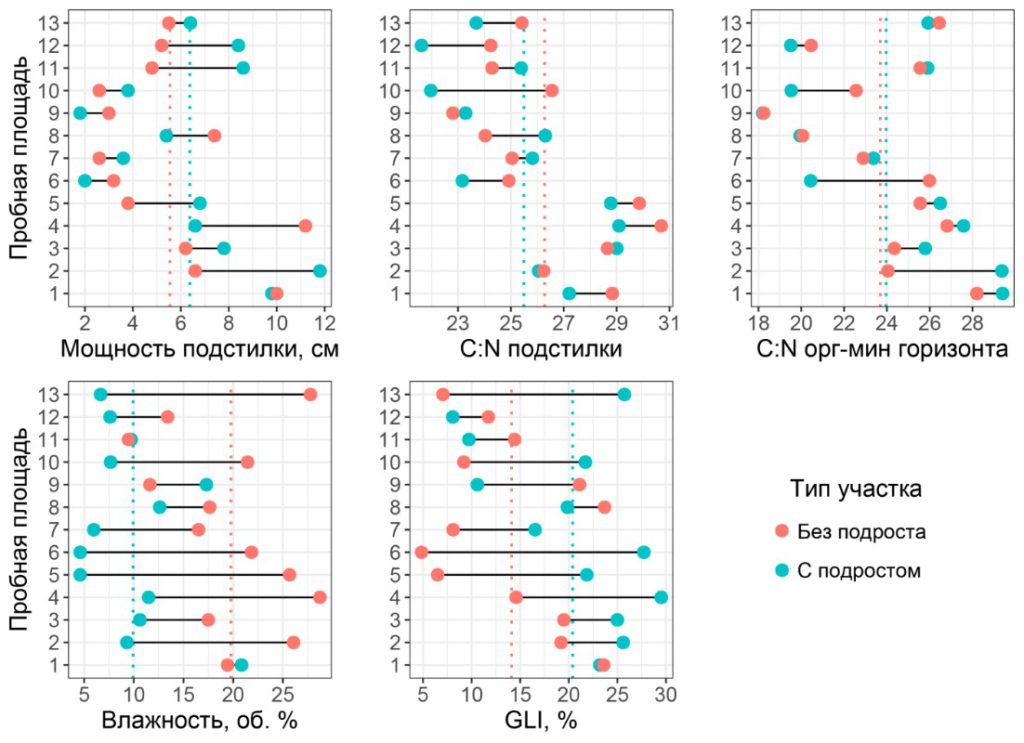
Рисунок 4. Значения исследованных экологических факторов в парах «контроль – опыт»
Примечание: пунктирными линиями показаны средние значения.
Взаимосвязь экологических факторов. Анализ комплекса экологических факторов показал наличие статистически значимых взаимосвязей между всеми исследованными почвенными характеристиками (рис. 5), кроме влажности и соотношения C:N органогенных и органо-минеральных горизонтов почвы; наиболее сильная взаимосвязь наблюдалась между соотношением C:N в органогенных и органо-минеральных горизонтах почвы.
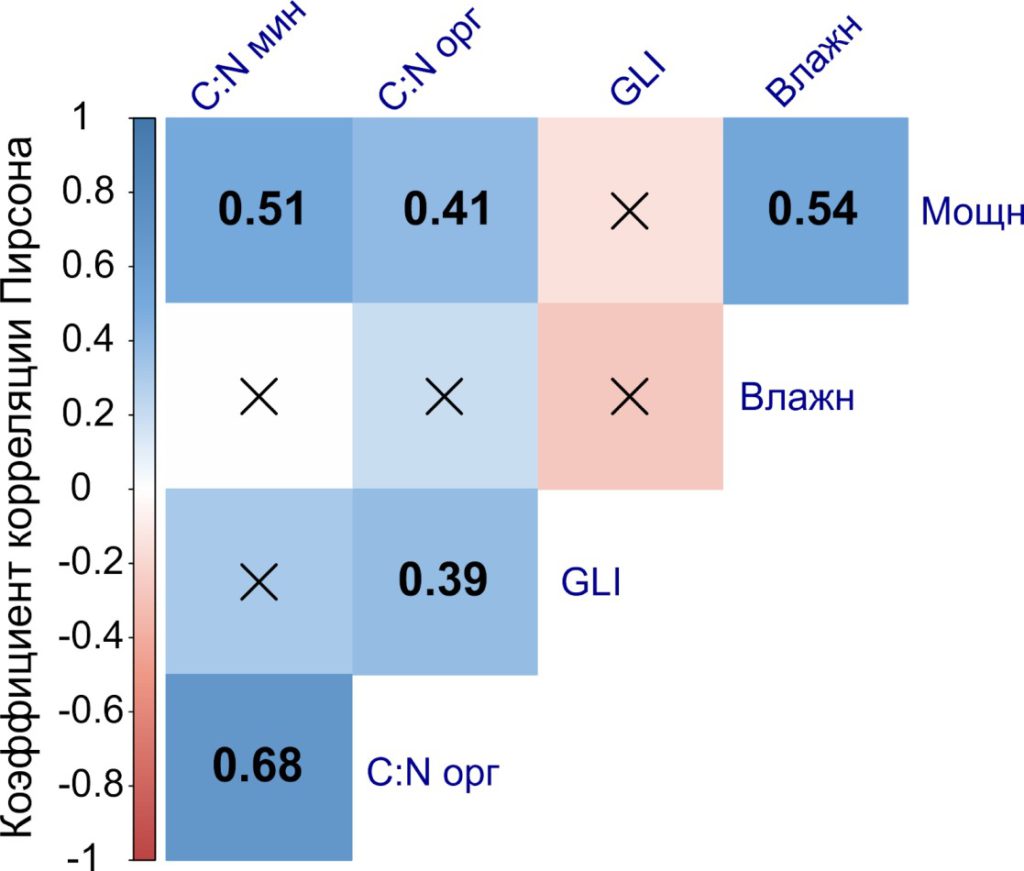
Рисунок 5. Коррелограмма взаимосвязей между исследованными экологическими факторами
Примечание: числами обозначены статистически значимые коэффициенты корреляции Пирсона (при уровне значимости 0.05)
Положительную корреляцию между влажностью и мощностью органогенных горизонтов можно объяснить тем, что более мощная подстилка замедляет процесс испарения влаги. Тесная взаимосвязь между соотношением C:N органогенных и органоминеральных горизонтов почвы объясняется тем, что в процессе гумификации, а также под действием почвенной фауны, опад постепенно переходит из органогенных в органо-минеральные горизонты и, таким образом, запасы углерода и азота в органо-минеральных горизонтах напрямую определяются его химическими свойствами. Положительная корреляция между мощностью подстилки и соотношением C:N в органогенных и органо-минеральных горизонтах может быть связана с более медленными процессами минерализации и гумификации опада, имеющего меньшую концентрацию азота, что в свою очередь приводит к накоплению подстилки.
Взаимосвязь между значениями GLI и различными характеристиками богатства почвы не была установлена, однако наблюдалась обратная линейная зависимость разности влажности почвы от разности GLI в парах «контроль – опыт» (рис. 6).
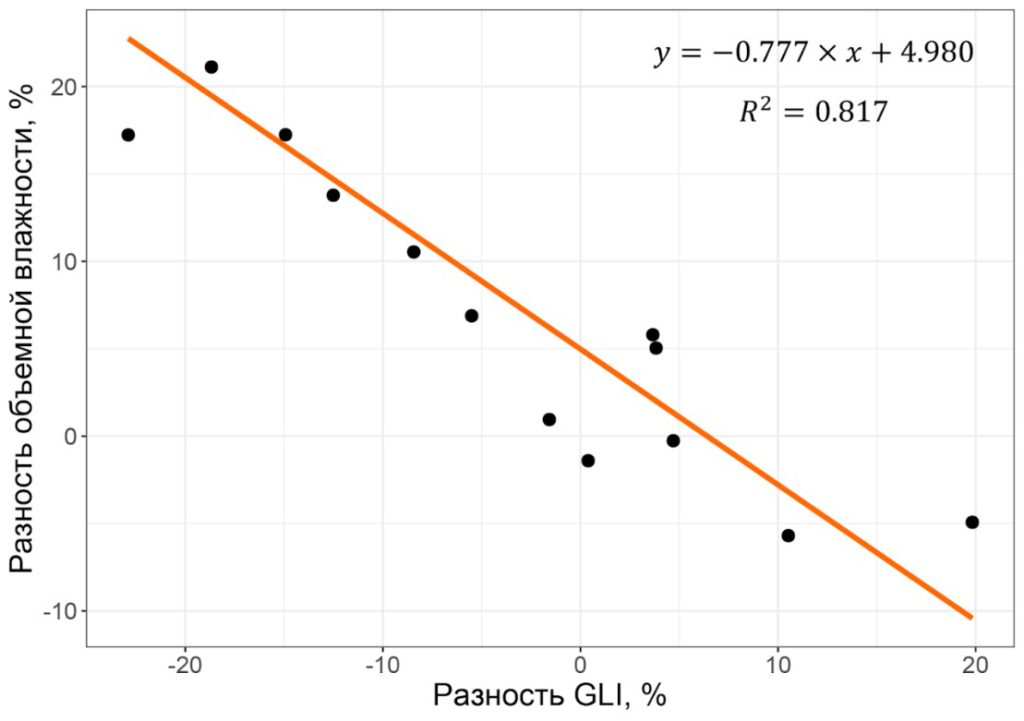
Рисунок 6. Зависимость разности объемной влажности органоминеральных горизонтов почвы от разности GLI в парах «контроль – опыт»
Наличие зависимости разности объемной влажности почвы в слое 0–6 см от разности GLI в парах «контроль–опыт» при отсутствии зависимости между абсолютными значениями этих факторов можно объяснить тем, что на влажность почвы, помимо инсоляции, оказывают влияние другие факторы, такие как микрорельеф, близость грунтовых вод и другие. При сравнении участков одной площадки с подростом и без него, измерения на которых проводились одновременно, влияние этих факторов было минимальным, и основным действующим фактором, влияющим на испарение влаги с поверхности почвы, и, как следствие, изменение объемной влажности в слое 0–6 см, являлась инсоляция. Это позволяет сказать, что, несмотря на статистическую значимость различий объемной влажности в парах «контроль–опыт», ключевую роль в наличии или отсутствии подроста среди абиотических факторов играет именно освещенность под пологом.
ЗАКЛЮЧЕНИЕ
В ходе проведенного исследования проанализированы зависимости жизненности и видового состава древесного подроста от комплекса биотических, абиотических и эдафических факторов. Нами не было выявлено зависимости возобновления деревьев от доминантов живого напочвенного покрова. Результаты исследования показали, что при всем многообразии и комплексности экологических факторов, влияющих на естественное лесовозобновление, можно установить их взаимозависимость и выделить ведущие. В исследованных сосновых сообществах такими факторами являются характеристики почвенного богатства и освещенность. При том, что видовой состав древесного подроста зависит от разных групп факторов (P. abies от почвенного богатства, а P. sylvestris – от влажности почвы и освещенности), жизненность подроста вне зависимости от вида, зависит в первую очередь от освещенности.
Анализ результатов позволяет количественно установить диапазоны толерантности видов по отношению к факторам среды и использовать результаты данного исследования при моделировании естественного лесовозобновления.
БЛАГОДАРНОСТИ
Работа выполнена при частичной финансовой поддержке РФФИ (грант 18-34-00556) в рамках тем государственного задания ИФХиБПП РАН № АААА-А18-118013190176-2, ИМПБ РАН – филиал ИПМ им. М.В. Келдыша РАН № 0017-2019-0009 и ЦЭПЛ РАН № АААА-А18-118052590019-7.
СПИСОК ЛИТЕРАТУРЫ
Алексеев В.А. Световой режим леса. Л.: Наука, 1975. 228 с.
Восточноевропейские леса: история в голоцене и современность. Отв. ред. О.В. Смирнова. М.: Наука, 2004. Книга 1. 479 с.
Гатцук Л.Е., Ермакова И.М. Общие представления о жизненном состоянии особи и ценопопуляции // Подходы к изучению ценопопуляций и консорций. М.: МГПИ, 1987. С. 3-7.
Грязькин А.В. Возобновительный потенциал таежных лесов (На примере ельников Северо-Запада России). СПб.: СПбГЛТА, 2001. 188 с.
Демаков Ю.П. Диагностика устойчивости лесных экосистем (методологические и методические аспекты). Йошкар-Ола: Периодика Марий Эл, 2000. 416 с.
Денисов С.А., Демичева Н.В., Егоров В.М. Теория и практика естественного возобновления леса. Режим доступа: http://csfm.volgatech.net/elearning/vozobnovlenie/text/ (дата обращения 15.07.2019).
Дружинин Н.А., Дружинин Ф.Н., Королева Н.С., Шушарин А.П. Лесовосстановительные процессы на торфяных почвах в естественных, осушаемых, пройденных рубками насаждениях // Лесопользование и гидролесомелиорация: материалы Всероссийского симпозиума. Ч. 1. СПб.; Вологда., 2007. С. 82-91.
Евстигнеев О.И. Световой минимум подроста деревьев хвойно-широколиственного леса // Структурно-функциональная организация и динамика растительного покрова. 2012. С. 181-184.
Жиляев Г.Г. Жизнеспособность популяций растений / отв. ред. К.А. Малиновский. Львов, 2005. 304 с.
Жукова Л.А. Многообразие путей онтогенеза в популяциях растений // Экология. 2001. № 3. С. 169-176.
Кобак К.И., Алексеев В.А. Световой и углекислотный режимы под пологом сосновых насаждений как условие произрастания подроcта // Световой режим, фотосинтез и продуктивность леса. Л., 1965. С. 23-24.
Комаров А.С., Припутина И.В., Михайлов А.В., Чертов О.Г. Биогеохимический цикл углерода в лесных экосистемах центра Европейской России и его техногенные изменения // Почвенные процессы и пространственно-временная организация почв. М.: Наука, 2006. С. 362-377.
Лесной план московской области на 2019-2028 годы: утвержден постановлением Губернатора Московской области от 21.03.2019 № 116-ПГ. Книга 1. 160 с.
Морозов Г.Ф. Учение о лесе. Вып. 1. Введение в биологию леса. СПб, 1912. 183 с.
Николаев Н.С., Аблеева В.А. Характеристика климатических показателей территории Приокско-террасного заповедника за период наблюдений 1973-2014 гг. // Труды Приокско-Террасного заповедника. Вып. 6. Тула: Аквариус, 2015. С. 11-23.
Рязапов Р.И., Кабанов С.В. Подпологовая освещенность в старовозрастных сосняках естественного происхождения южной части Приволжской возвышенности и ее влияние на жизненность подроста сосны // Вестник Алтайского государственного аграрного университета. 2011. Т. 77. №. 3. С. 54-60.
Санников С.Н., Санникова Н.С. Экология естественного возобновления сосны под пологом леса. М.: Наука, 1985. 152 с.
Санников С.Н., Санникова Н.С. Лес как подземно-сомкнутая дендроценоэкосистема // Сибирский лесной журнал. 2014. №. 1. С. 25-34.
Смирнов Е.Г., Беленец Ю.Е. Состояние лесовосстановления в условиях черничниковой группы типов леса Ленинградской области // Лесопользование и гидролесомелиорация: материалы Всероссийского симпозиума. СПб.; Вологда: СевНИИЛХ, 2007. С. 76-78
Суслова Е.Г. Леса Московской области // Экосистемы: экология и динамика. 2019. Т. 3. №. 1. с. 119-190
Уткин А.И., Замолодчиков Д.Г., Честных О.В., Коровин Г.Н., Зукерт Н.В. Леса России как резервуар органического углерода биосферы // Лесоведение. 2001. № 5. С. 8-23.
Цветков В.Ф. Проблемы лесовозобновления на европейском севере России. Лесные ресурсы таежной зоны России: проблемы лесопользования и лесовосстановления // Материалы Всероссийской научной конференции с международным участием. Петрозаводск: КарНЦ РАН, 2009. С. 153-156.
Чучалина А.А., Мищихина Ю.Д. Численность, возрастная структура и жизненность подроста сосны в контрастных экотопах подзон предлесостепи и средней тайги // Известия Оренбургского государственного аграрного университета. 2013. №. 1. С. 14-17
Шаньгина Н.П., Феклистов П.А. Возобновление коренных ельников и влияние экологических факторов на формирование подроста под пологом древостоев // Проблемы экологии в современном мире в свете учения В.И. Вернадского: материалы межд. конф. Тамбов.: Дом ТГУ, 2010. Т. 2. С.118-122.
Экологический мониторинг Серпуховского района. Отчет о работе по договору о научно-техническом сотрудничестве между ИПФС АН СССР и ПТЗ. 1989.
Aas G. Quercus petraea (Mattuschka) Lieblein, 1784. Traubeneiche. In: Roloff A, Weisgerber H, Lang U, Stimm B, Schütt P, editors. Enzyklopädie der Holzgewächse. Handbuch und Atlas der Dendrologie: Wiley-VCH, 2002. P. 1-16.
Aas G. Quercus robur L., 1753. Stieleiche. In: Roloff A, Weisgerber H, Lang U, Stimm B, Schütt P, editors. Enzyklopädie der Holzgewächse. Handbuch und Atlas der Dendrologie: Wiley-VCH, 2002. P. 17-29.
Annighöfer P., Beckschäfer P., Vor T., Ammer C. Regeneration patterns of European oak species (Quercus petraea (Matt.) Liebl., Quercus robur L.) in dependence of environment and neighborhood // PloS one. 2015. Vol. 10. No. 8. P. 1-16.
Axelsson E., Lundmark T., Högberg P., Nordin A. Belowground competition directs spatial patterns of seedling growth in boreal pine forests in Fennoscandia // Forests. 2014. Vol. 5. No. 9. P. 2106-2121.
Beland M., Agestam E., Ekö P.M., Gemmel P., Nilsson U. Scarification and seedfall affects natural regeneration of Scots pine under two shelterwood densities and a clear-cut in southern Sweden // Scandinavian Journal of Forest Research. 2000. Vol. 15. No. 2. P. 247-255.
Booth T.C. Natural regeneration in the nativepinewoods of Scotland. A review of principles and practice // Scottish Forestry. 1984. Vol 38. P. 33-42.
Čater M., Batič F. Groundwater and light conditions as factors in the survival of pedunculate oak (Quercus robur L.) seedlings // European Journal of Forest Research. 2006. Vol. 125. No. 4. P. 419-426.
Caudullo G., Tinner W., de Rigo D. Picea abies in Europe: distribution, habitat, usage and threats // European Atlas of Forest Tree Species. San-Miguel-Ayanz J., de Rigo D., Caudullo G., Houston Durrant T.; Mauri A. (eds.). 2016. P. 114-116.
Cescatti A. Indirect estimates of canopy gap fraction based on the linear conversion of hemispherical photographs: Methodology and comparison with standard thresholding techniques // Agricultural and Forest Meteorology. 2007. Vol. 143. No. 1-2. P. 1-12.
Chulliat A., Macmillan S., Alken P., Beggan C., Nair M., Hamilton B., Woods A., Ridley V., Maus S., Thomson A. The US/UK World Magnetic Model for 2015-2020: Technical Report, National Geophysical Data Center, NOAA, 2015.
Coban S., Colak A.H., Rotherham I.D. Interactions between canopy cover density and regeneration cores of older saplings in Scots pine (Pinus sylvestris L.) stands // Forest systems. 2016. Vol. 25. No. 3. P. 1-14.
Dyderski M.K., Paź S., Frelich L.E., Jagodziński A.M. How much does climate change threaten European forest tree species distributions? // Global Change Biology. 2018. Vol. 24. No. 3. P. 1150-1163.
Ellenberg H., Leuschner C. Vegetation Mitteleuropas mit den Alpen: in ökologischer, dynamischer und historischer Sicht. Utb, 2010. Vol. 8104. 1095 p.
Evstigneev O.I. Ontogenetic scales of relation of trees to light (on the example of Eastern European forests) // Russian Journal of Ecosystem Ecology. 2018. No. 3. P. 1-18.
Gill R.M.A. A review of damage by mammals in north temperate forests: 3. Impact on trees and forests // Forestry: An International Journal of Forest Research. 1992. Vol. 65. No. 4. P. 363-388.
Gong Y. Studies on the regeneration of Pinus sylvestris and Betula pendula: Diss. University of Aberdeen, 1992. 234 p.
Gutowski J.M., Jaroszewicz B. Zmiany udziału świerka pospolitego w drzewostanach Puszczy Białowieskiej w kontekście dynamiki liczebności kornika drukarza // Wikło A (ed.) Stan ekosystemów leśnych Puszczy Białowieskiej. 2015. P. 87-108.
Jarčuška B., Kucbel S., Jaloviar P. Comparison of output results from two programmes for hemispherical image analysis: Gap Light Analyser and WinScanopy // Journal of Forest Science. 2010. Vol. 56. No. 4. P. 147-153.
Jonckheere I., Fleck S., Nackaerts K., Muys B., Coppin P., Weiss M., Baret F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography // Agricultural and Forest Meteorology. 2004. Vol. 121. No. 1-2. P. 19-35.
Jonckheere I., Nackaerts K., Muys B., Coppin P. Assessment of automatic gap fraction estimation of forests from digital hemispherical photography // Agricultural and Forest Meteorology. 2005. Vol. 132. No. 1-2. P. 96-114.
Kielland-Lund J. Die Waldgesellschaften SO-Norwegens // Phytocoenologia. 1981. P. 53-250.
Kuuluvainen T., Ylläsjärvi I. On the natural regeneration of dry heath forests in Finnish Lapland: a review of VT Aaltonen (1919) // Scandinavian Journal of Forest Research. 2011. Vol. 26. No. S10. P. 34-44.
Leblanc S.G., Chen J.M., Fernandes R., Deering D.W., Conley A. Methodology comparison for canopy structure parameters extraction from digital hemispherical photography in boreal forests // Agricultural and Forest Meteorology. 2005. Vol. 129. No. 3-4. P. 187-207.
Macfarlane C., Coote M., White D.A., Adams M.A. Photographic exposure affects indirect estimation of leaf area in plantations of Eucalyptus globulus Labill // Agricultural and Forest Meteorology. 2000. Vol. 100. No. 2-3. P. 155-168.
Miina J., Saksa T. Predicting establishment of tree seedlings in regeneration areas of Picea abies in southern Finland // Baltic Forestry. 2013. Vol. 19. No. 2. P. 187-200.
Montes F., Cañellas I. The spatial relationship between post‐crop remaining trees and the establishment of saplings in Pinus sylvestris stands in Spain // Applied Vegetation Science. 2007. Vol. 10. No. 2. P. 151-160.
Niinemets Ü., Valladares F. Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs // Ecological Monographs. 2006. Vol. 76. No. 4. P. 521-547.
Nilsson U., Gemmel P., Löf M., Welander T. Germination and early growth of sown Quercus robur L. in relation to soil preparation, sowing depths and prevention against predation // New Forests. 1996. Vol.12. P. 69-86.
Nobis M., Hunziker U. Automatic thresholding for hemispherical canopy-photographs based on edge detection // Agricultural and Forest Meteorology. 2005. Vol. 128. No. 3-4. P. 243-250.
Poorter H., Niklas K.J., Reich P.B., Oleksyn J., Poot P., Mommer L. Biomass allocation to leaves, stems and roots: meta‐analyses of interspecific variation and environmental control // New Phytologist. 2012. Vol. 193. No. 1. P. 30-50.
Poorter H., De Jong R.O.B. A comparison of specific leaf area, chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity // New Phytologist. 1999. Vol. 143. No. 1. P. 163-176.
R Core Team R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2019. URL: https://www. R-project. org (aссessed October 31, 2019).
Rich P.M. Characterizing plant canopies with hemispherical photographs // Remote Sensing Reviews. 1990. Vol. 5. No. 1. P. 13-29.
Ruuska J., Siipilehto J., Valkonen S. Effect of edge stands on the development of young Pinus sylvestris stands in southern Finland // Scandinavian Journal of Forest Research. 2008. Vol. 23. No. 3. P. 214-226.
Siipilethto J. Height distributions of Scots pine sapling stands affected by retained tree and edge stand competition // Silva Fennica 2006. Vol. 40. No 3. P. 473-486.
Šúri M., Huld T.A., Dunlop E.D. PV-GIS: a web-based solar radiation database for the calculation of PV potential in Europe // International Journal of Sustainable Energy. 2005. Vol. 24. No. 2. P. 55-67.
Szydlarski M., Modrzyński J. Increase of natural regeneration area of Norway spruce (Picea abies L. Karst.) in the Kaszuby Lake District during the decade 2002-2012 // Forest Research Papers. 2015. Vol. 76. No. 1. P. 66-72.
Thimonier A., Sedivy I., Schleppi P. Estimating leaf area index in different types of mature forest stands in Switzerland: a comparison of methods // European Journal of Forest Research. 2010. Vol. 129. No. 4. P. 543-562.
Wagner S. Calibration of grey values of hemispherical photographs for image analysis / /Agricultural and Forest Meteorology. 1998. Vol. 90. No. 1-2. P. 103-117.
Wittich W. Die standörtlichen Bedingungen für die natürliche Verjüngung der Kiefer und für ihre Erziehung unter Schirm // Allgemeine Forst-und Jagdzeitung. 1955. Vol. 126. P. 109-117.
Wyka T.P., Oleksyn J., Żytkowiak R., Karolewski P., Jagodziński A.M., Reich P.B. Responses of leaf structure and photosynthetic properties to intra-canopy light gradients: a common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species // Oecologia. 2012. Vol. 170. No. 1. P. 11-24.
Yazdani R., Lindgren D. Hur tallens gener sprids vid naturlig föryngring under fröträd. // SLU. Skogsfakta. Biologi och skogsskötsel. 1990. 74 p.
Zhang Y., Chen J.M., Miller J.R. Determining digital hemispherical photograph exposure for leaf area index estimation // Agricultural and Forest Meteorology. 2005. Vol. 133. No. 1-4. P. 166-181.
Рецензент: к.б.н., в.н.с. Тихонова Е.В.





